

Analysis of mutations from SCID and Omenn syndrome patients reveals the central role of the Rag2 PHD domain in regulating V(D)J recombination
- PMID: 20234091
- PMCID: PMC2846059
- DOI: 10.1172/JCI41305
Analysis of mutations from SCID and Omenn syndrome patients reveals the central role of the Rag2 PHD domain in regulating V(D)J recombination
Abstract
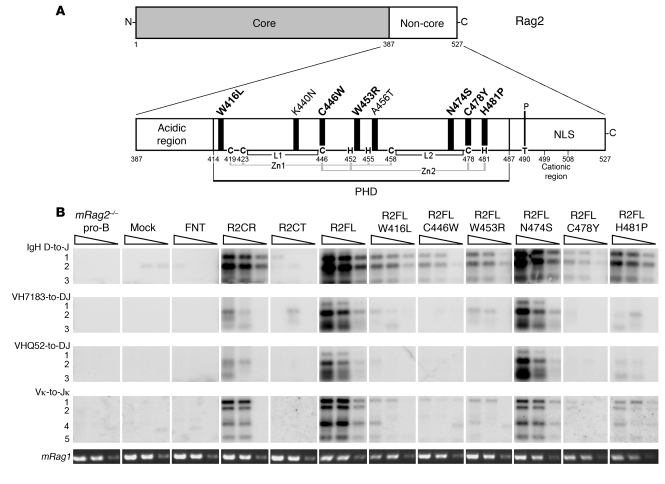
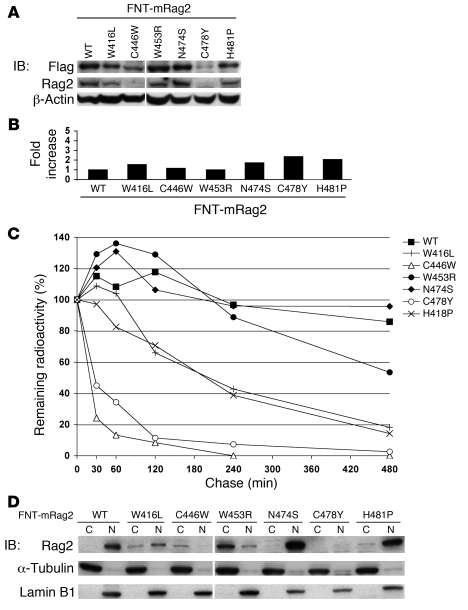
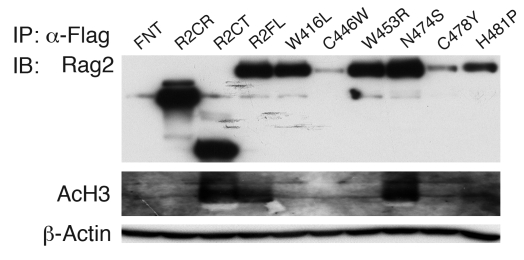
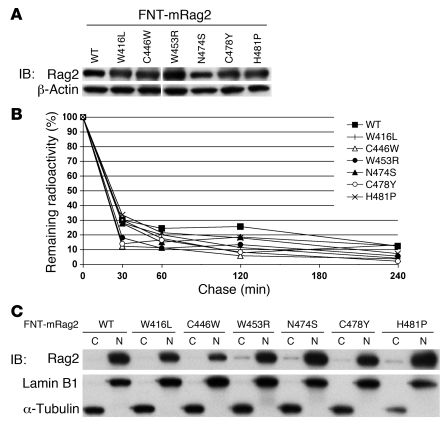
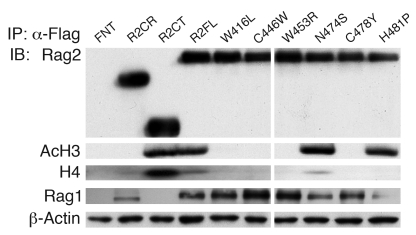
Rag2 plays an essential role in the generation of antigen receptors. Mutations that impair Rag2 function can lead to severe combined immunodeficiency (SCID), a condition characterized by complete absence of T and B cells, or Omenn syndrome (OS), a form of SCID characterized by the virtual absence of B cells and the presence of oligoclonal autoreactive T cells. Here, we present a comparative study of a panel of mutations that were identified in the noncanonical plant homeodomain (PHD) of Rag2 in patients with SCID or OS. We show that PHD mutant mouse Rag2 proteins that correspond to those found in these patients greatly impaired endogenous recombination of Ig gene segments in a Rag2-deficient pro-B cell line and that this correlated with decreased protein stability, impaired nuclear localization, and/or loss of the interaction between Rag2 and core histones. Our results demonstrate that point mutations in the PHD of Rag2 compromise the functionality of the entire protein, thus explaining why the phenotype of cells expressing PHD point mutants differs from those expressing core Rag2 protein that lacks the entire C-terminal region and is therefore devoid of the regulation imposed by the PHD. Together, our findings reveal the various deleterious effects of PHD Rag2 mutations and demonstrate the crucial role of this domain in regulating antigen receptor gene assembly. We believe these results reveal new mechanisms of immunodeficiency in SCID and OS.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
