

RPL41, a small ribosomal peptide deregulated in tumors, is essential for mitosis and centrosome integrity
- PMID: 20234822
- PMCID: PMC2838445
- DOI: 10.1593/neo.91610
RPL41, a small ribosomal peptide deregulated in tumors, is essential for mitosis and centrosome integrity
Abstract
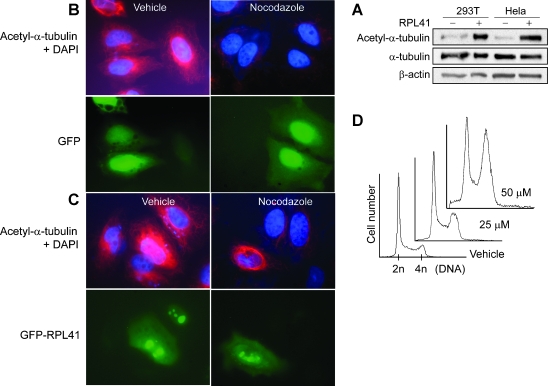
Ribosomal large subunit protein RPL41 is a basic (positively charged) peptide consisting of only 25 amino acids. An antisense-based functional screening revealed that the down-regulation of RPL41 led to an anchorage-independent growth of NIH3T3 cells in soft agar plates. RPL41 depletion with gene-specific small interfering RNA also resulted in malignant transformation of NIH3T3 cells including increased tumor growth in mice. RPL41 deletion was detected in 59% of tumor cell lines by fluorescence in situ hybridization analyses and RPL41 down-regulation in 75% of primary breast cancers by real-time quantitative reverse transcription-polymerase chain reaction. These studies suggest a tumor suppression role for RPL41. By mass spectrometry, RPL41 was associated with several cytoskeleton components including tubulin beta, gamma, and myosin IIA, which was confirmed by Western blot analysis on both cellular lysis and individually in vitro-expressed proteins. RPL41 also bound directly to polymerized tubulins. Cells overexpressing a GFP-RPL41 were resistant to nocodazole-induced microtubule depolymerization. A synthetic RPL41 induced cellular alpha-tubulin acetylation and G(2)/M cell cycle arrest. These results indicate a stabilizing role of RPL41 on microtubule. Microtubule spindles are essential for chromosome segregation during mitosis. Cells with RPL41 knock-down showed abnormal spindles, frequent failure of cytokinesis, and formation of polynuclear cells. In interphase cells, RPL41-depleted cells had premature splitting of centrosome. Our results provide evidence that RPL41 is a microtubule-associated protein essential for functional spindles and for the integrity of centrosome and that the abnormal mitosis and disrupted centrosome associated with the RPL41 down-regulation may be related to malignant transformation.
Figures






Similar articles
-
Gamma-actin is involved in regulating centrosome function and mitotic progression in cancer cells.Cell Cycle. 2015;14(24):3908-19. doi: 10.1080/15384101.2015.1120920. Cell Cycle. 2015. PMID: 26697841 Free PMC article.
-
The sudden recruitment of gamma-tubulin to the centrosome at the onset of mitosis and its dynamic exchange throughout the cell cycle, do not require microtubules.J Cell Biol. 1999 Aug 9;146(3):585-96. doi: 10.1083/jcb.146.3.585. J Cell Biol. 1999. PMID: 10444067 Free PMC article.
-
A centrosomal protein STARD9 promotes microtubule stability and regulates spindle microtubule dynamics.Cell Cycle. 2018;17(16):2052-2068. doi: 10.1080/15384101.2018.1513764. Epub 2018 Sep 11. Cell Cycle. 2018. PMID: 30160609 Free PMC article.
-
Mitotic spindle multipolarity without centrosome amplification.Nat Cell Biol. 2014 May;16(5):386-94. doi: 10.1038/ncb2958. Nat Cell Biol. 2014. PMID: 24914434 Review.
-
Positioning centrosomes and spindle poles: looking at the periphery to find the centre.Biol Cell. 2006 Sep;98(9):557-65. doi: 10.1042/BC20060017. Biol Cell. 2006. PMID: 16907664 Review.
Cited by
-
The lncRNA ADAMTS9-AS2 Regulates RPL22 to Modulate TNBC Progression via Controlling the TGF-β Signaling Pathway.Front Oncol. 2021 Jun 9;11:654472. doi: 10.3389/fonc.2021.654472. eCollection 2021. Front Oncol. 2021. PMID: 34178640 Free PMC article.
-
Overcoming intratumor heterogeneity of polygenic cancer drug resistance with improved biomarker integration.Neoplasia. 2012 Dec;14(12):1278-89. doi: 10.1593/neo.122096. Neoplasia. 2012. PMID: 23308059 Free PMC article.
-
The interconnectedness of cancer cell signaling.Neoplasia. 2011 Dec;13(12):1183-93. doi: 10.1593/neo.111746. Neoplasia. 2011. PMID: 22241964 Free PMC article.
-
RNA-Protein Interactome at the Hepatitis E Virus Internal Ribosome Entry Site.Microbiol Spectr. 2023 Aug 17;11(4):e0282722. doi: 10.1128/spectrum.02827-22. Epub 2023 Jun 29. Microbiol Spectr. 2023. PMID: 37382527 Free PMC article.
-
Ribosomal protein S27-like regulates autophagy via the β-TrCP-DEPTOR-mTORC1 axis.Cell Death Dis. 2018 Nov 13;9(11):1131. doi: 10.1038/s41419-018-1168-7. Cell Death Dis. 2018. PMID: 30425236 Free PMC article.
References
-
- Zimmermann RA. The double life of ribosomal proteins. Cell. 2003;115:130–132. - PubMed
-
- Mazumder B, Sampath P, Seshadri V, Maitra RK, DiCorleto PE, Fox PL. Regulated release of L13a from the 60S ribosomal subunit as a mechanism of transcript-specific translational control. Cell. 2003;115:187–198. - PubMed
-
- Chavez-Rios R, Arias-Romero LE, Almaraz-Barrera MJ, Hernandez-Rivas R, Guillen N, Vargas M. L10 ribosomal protein from Entamoeba histolytica share structural and functional homologies with QM/Jif-1: proteins with extraribosomal functions. Mol Biochem Parasitol. 2003;127:151–160. - PubMed
-
- Neumann F, Krawinkel U. Constitutive expression of human ribosomal protein L7 arrests the cell cycle in G1 and induces apoptosis in Jurkat T-lymphoma cells. Exp Cell Res. 1997;230:252–261. - PubMed
-
- Khanna N, Reddy VG, Tuteja N, Singh N. Differential gene expression in apoptosis: identification of ribosomal protein S29 as an apoptotic inducer. Biochem Biophys Res Commun. 2000;277:476–486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials