

Lymphangioleiomyomatosis and TSC2-/- cells
- PMID: 20235888
- PMCID: PMC2883471
- DOI: 10.1089/lrb.2009.0031
Lymphangioleiomyomatosis and TSC2-/- cells
Abstract
The cells comprising pulmonary lymphangioleiomyomatosis (LAM) and renal angiomyolipomas (AMLs) are heterogeneous, with variable mixtures of cells exhibiting differentiation towards smooth muscle, fat, and vessels. Cells grown from LAM and AMLs have likewise tended to be heterogeneous. The discovery that LAM and AMLs contain cells with mutations in the TSC1 or TSC2 genes is allowing investigators to discriminate between "two-hit" cells and neighboring cells, providing insights into disease pathogenesis. In rare cases, it has been possible to derive cells from human tumors, including AMLs and TSC skin tumors that are highly enriched for TSC2(-/-) cells. Cells derived from an Eker rat uterine leiomyoma (ELT3 cells) are Tsc2-null and these have been used in a rodent cell models for LAM. Further improvements in the ability to reliably grow well-characterized TSC2(-/-) cells from human tumors are critical to developing in vitro and in vivo model systems for studies of LAM pathogenesis and treatment.
Figures
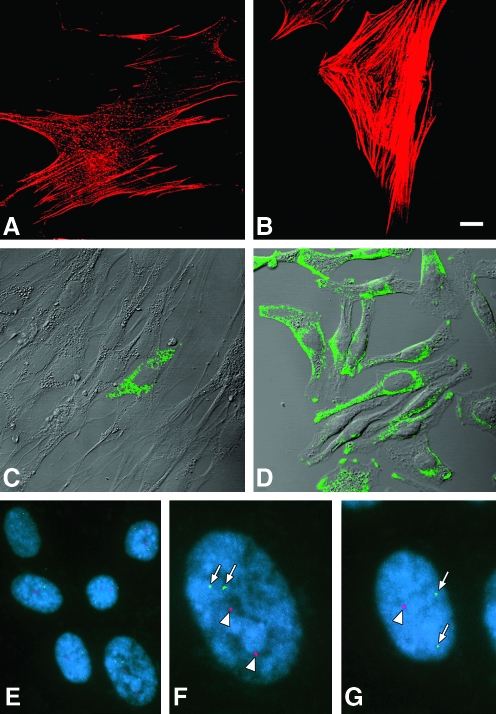
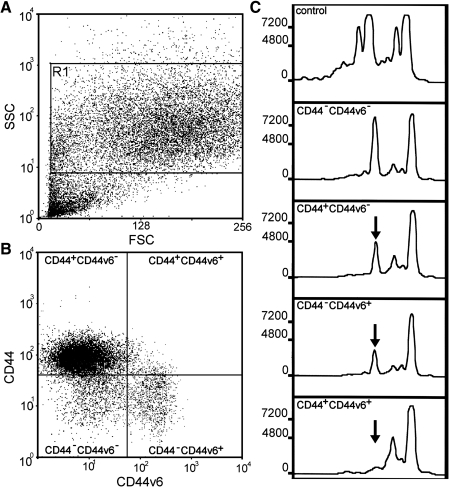
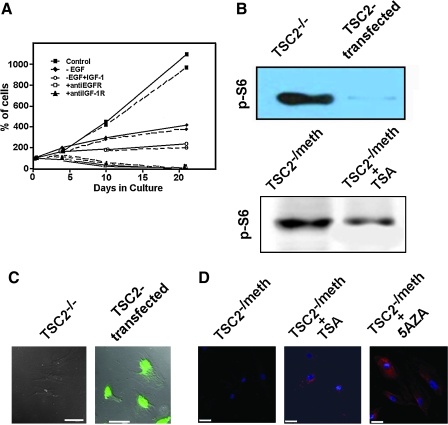
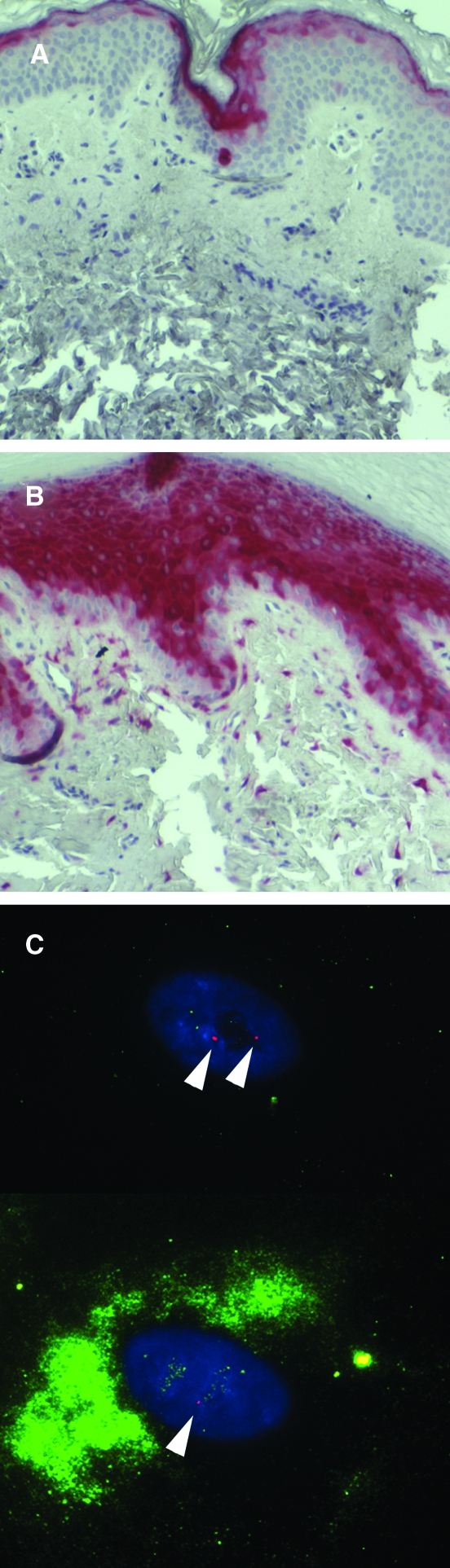
Similar articles
-
Rapamycin-insensitive up-regulation of adipocyte phospholipase A2 in tuberous sclerosis and lymphangioleiomyomatosis.PLoS One. 2014 Oct 27;9(10):e104809. doi: 10.1371/journal.pone.0104809. eCollection 2014. PLoS One. 2014. PMID: 25347447 Free PMC article.
-
Animal models of lymphangioleiomyomatosis (LAM) and tuberous sclerosis complex (TSC).Lymphat Res Biol. 2010 Mar;8(1):51-7. doi: 10.1089/lrb.2009.0013. Lymphat Res Biol. 2010. PMID: 20235887 Free PMC article. Review.
-
Interferon beta augments tuberous sclerosis complex 2 (TSC2)-dependent inhibition of TSC2-null ELT3 and human lymphangioleiomyomatosis-derived cell proliferation.Mol Pharmacol. 2008 Mar;73(3):778-88. doi: 10.1124/mol.107.040824. Epub 2007 Dec 19. Mol Pharmacol. 2008. PMID: 18094073
-
Estrogen maintains myometrial tumors in a lymphangioleiomyomatosis model.Endocr Relat Cancer. 2016 Apr;23(4):265-80. doi: 10.1530/ERC-15-0505. Epub 2016 Feb 15. Endocr Relat Cancer. 2016. PMID: 26880751 Free PMC article.
-
Pathogenesis of multifocal micronodular pneumocyte hyperplasia and lymphangioleiomyomatosis in tuberous sclerosis and association with tuberous sclerosis genes TSC1 and TSC2.Pathol Int. 2001 Aug;51(8):585-94. doi: 10.1046/j.1440-1827.2001.01242.x. Pathol Int. 2001. PMID: 11564212 Review.
Cited by
-
Diagnostic value of serum vascular endothelial growth factor-D in Korean patients with lymphangioleiomyomatosis.Ther Adv Respir Dis. 2024 Jan-Dec;18:17534666241272928. doi: 10.1177/17534666241272928. Ther Adv Respir Dis. 2024. PMID: 39148439 Free PMC article.
-
High Mobility Group AT-Hook 2 (HMGA2) Oncogenicity in Mesenchymal and Epithelial Neoplasia.Int J Mol Sci. 2020 Apr 29;21(9):3151. doi: 10.3390/ijms21093151. Int J Mol Sci. 2020. PMID: 32365712 Free PMC article. Review.
-
Treatment option(s) for pulmonary lymphangioleiomyomatosis: progress and current challenges.Am J Respir Cell Mol Biol. 2012 May;46(5):563-5. doi: 10.1165/rcmb.2011-0381ED. Am J Respir Cell Mol Biol. 2012. PMID: 22550272 Free PMC article. No abstract available.
-
Thoracoabdominal imaging of tuberous sclerosis.Pediatr Radiol. 2018 Aug;48(9):1307-1323. doi: 10.1007/s00247-018-4123-y. Epub 2018 Aug 4. Pediatr Radiol. 2018. PMID: 30078036 Review.
-
Evidence Supporting a Lymphatic Endothelium Origin for Angiomyolipoma, a TSC2(-) Tumor Related to Lymphangioleiomyomatosis.Am J Pathol. 2016 Jul;186(7):1825-1836. doi: 10.1016/j.ajpath.2016.03.009. Epub 2016 Jun 8. Am J Pathol. 2016. PMID: 27289491 Free PMC article.
References
-
- Taveira–DaSilva AM. Steagall WK. Moss J. Lymphangioleiomyomatosis. Cancer Control. 2006;13:276–285. - PubMed
-
- Zhe X. Schuger L. Combined smooth muscle and melanocytic differentiation in lymphangioleiomyomatosis. J Histochem Cytochem. 2004;52:1537–1542. - PubMed
-
- Folpe AL. Kwiatkowski DJ. Perivascular epithelioid cell neoplasms: Pathology and pathogenesis. Hum Pathol. 2010;41:1–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
