

USF and NF-E2 cooperate to regulate the recruitment and activity of RNA polymerase II in the beta-globin gene locus
- PMID: 20236933
- PMCID: PMC2871457
- DOI: 10.1074/jbc.M109.098376
USF and NF-E2 cooperate to regulate the recruitment and activity of RNA polymerase II in the beta-globin gene locus
Abstract
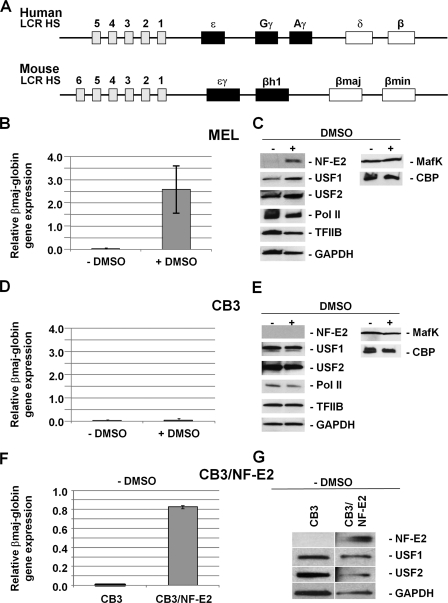
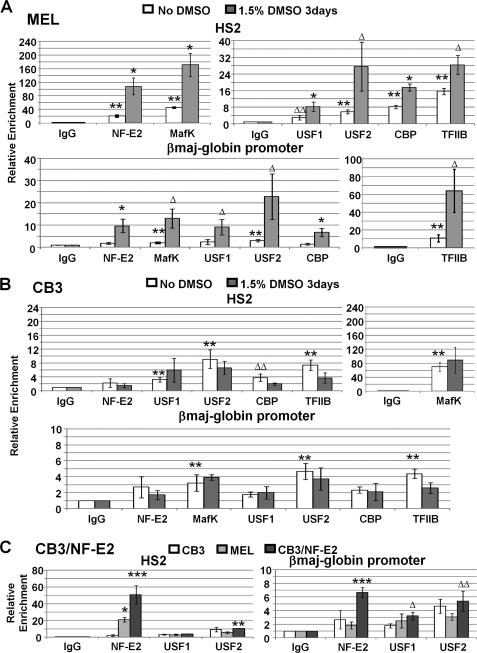
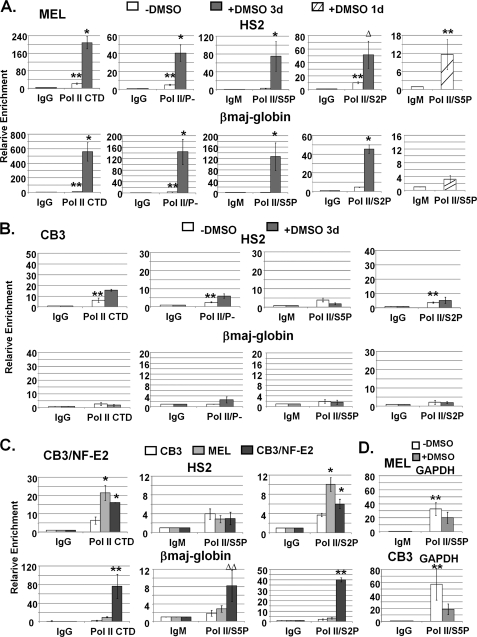
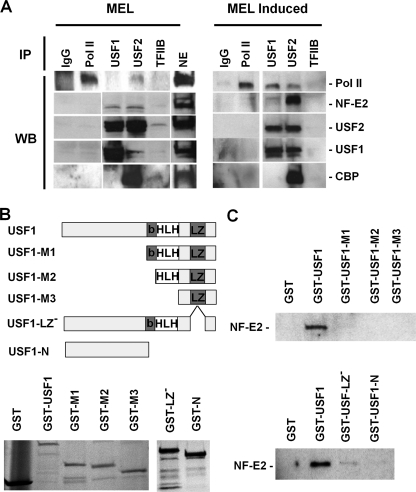
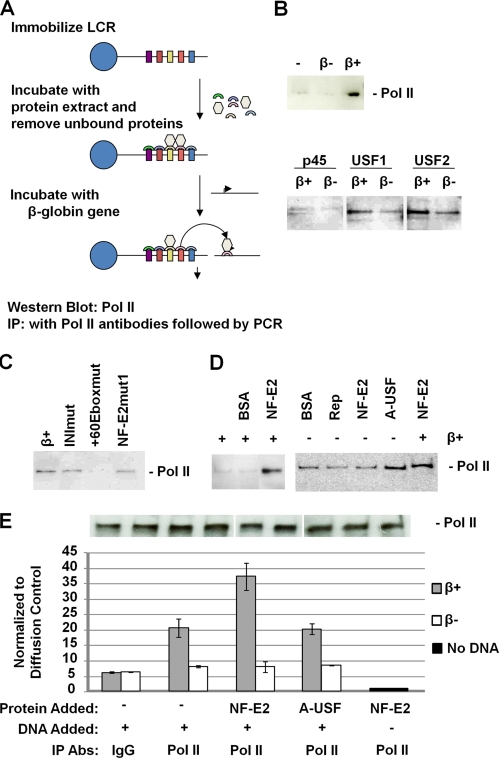
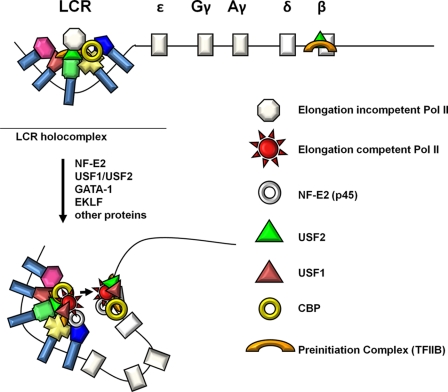
The human beta-globin gene is expressed at high levels in erythroid cells and regulated by proximal and distal cis-acting DNA elements, including promoter, enhancer, and a locus control region (LCR). Transcription complexes are recruited not only to the globin gene promoters but also to the LCR. Previous studies have implicated the ubiquitously expressed transcription factor USF and the tissue-restricted activator NF-E2 in the recruitment of transcription complexes to the beta-globin gene locus. Here we demonstrate that although USF is required for the efficient association of RNA polymerase II (Pol II) with immobilized LCR templates, USF and NF-E2 together regulate the association of Pol II with the adult beta-globin gene promoter. Recruitment of Pol II to the LCR occurs in undifferentiated murine erythroleukemia cells, but phosphorylation of LCR-associated Pol II at serine 5 of the C-terminal domain is mediated by erythroid differentiation and requires the activity of NF-E2. Furthermore, we provide evidence showing that USF interacts with NF-E2 in erythroid cells. The data provide mechanistic insight into how ubiquitous and tissue-restricted transcription factors cooperate to regulate the recruitment and activity of transcription complexes in a tissue-specific chromatin domain.
Figures
References
-
- Felsenfeld G., Groudine M. (2003) Nature 421, 448–453 - PubMed
-
- Chakalova L., Debrand E., Mitchell J. A., Osborne C. S., Fraser P. (2005) Nat. Rev. Genet. 6, 669–677 - PubMed
-
- Sutherland H., Bickmore W. A. (2009) Nat. Rev. Genet. 10, 457–466 - PubMed
-
- Loose M., Swiers G., Patient R. (2007) Curr. Opin. Hematol. 14, 307–314 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
