

FNR-mediated regulation of bioluminescence and anaerobic respiration in the light-organ symbiont Vibrio fischeri
- PMID: 20298504
- PMCID: PMC2866068
- DOI: 10.1111/j.1574-6968.2010.01938.x
FNR-mediated regulation of bioluminescence and anaerobic respiration in the light-organ symbiont Vibrio fischeri
Abstract
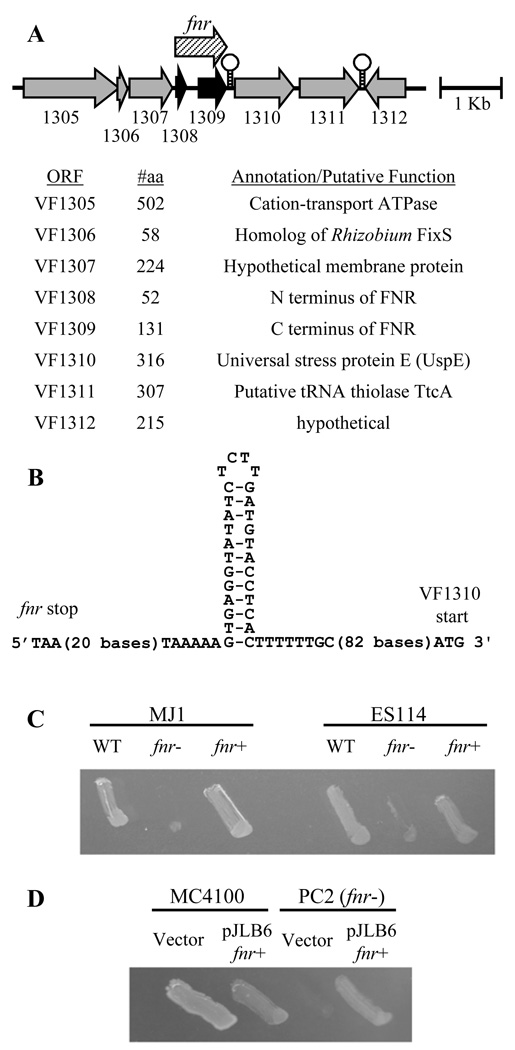
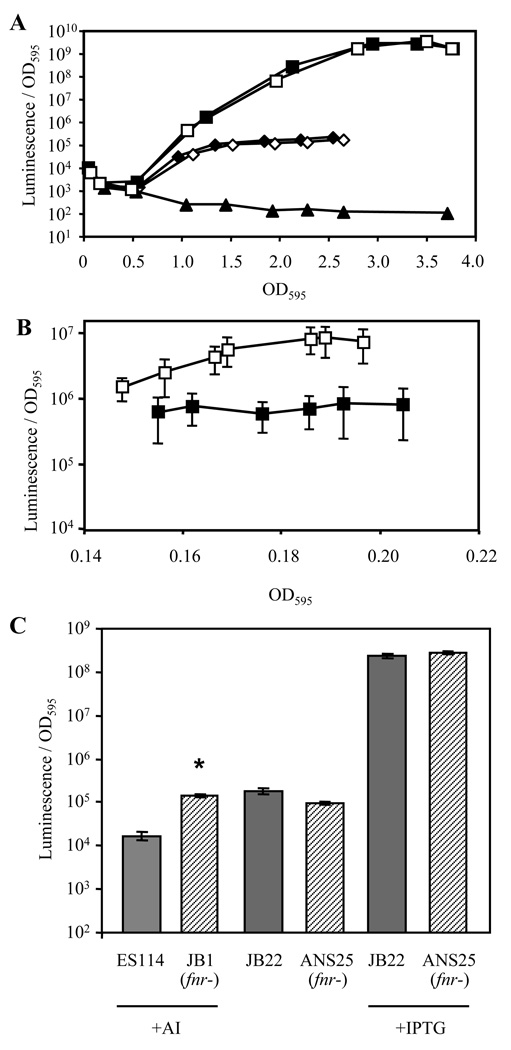
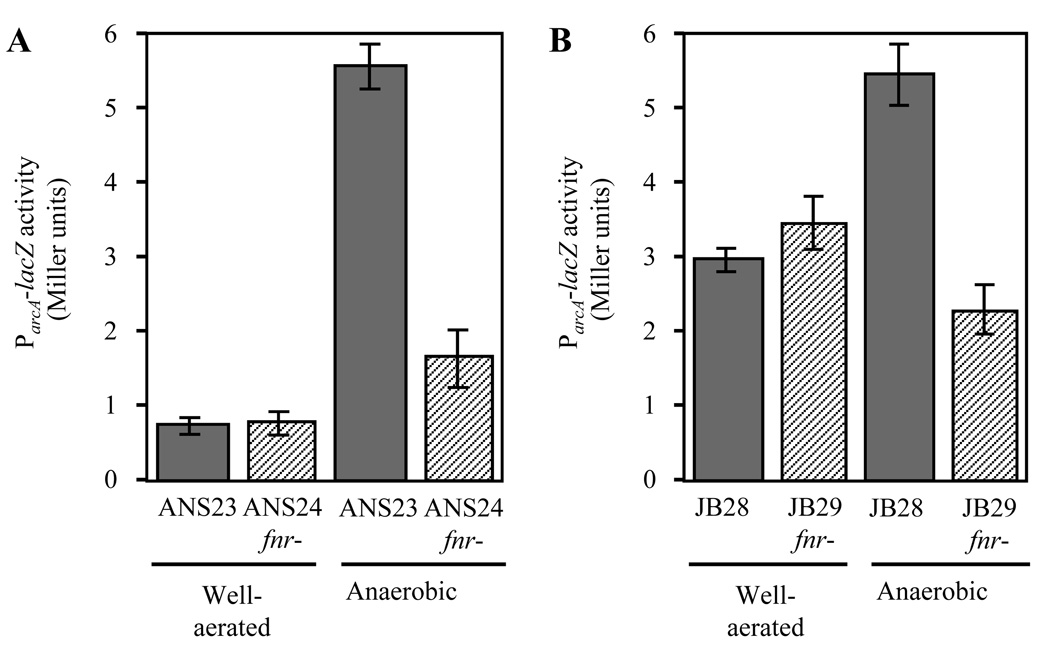
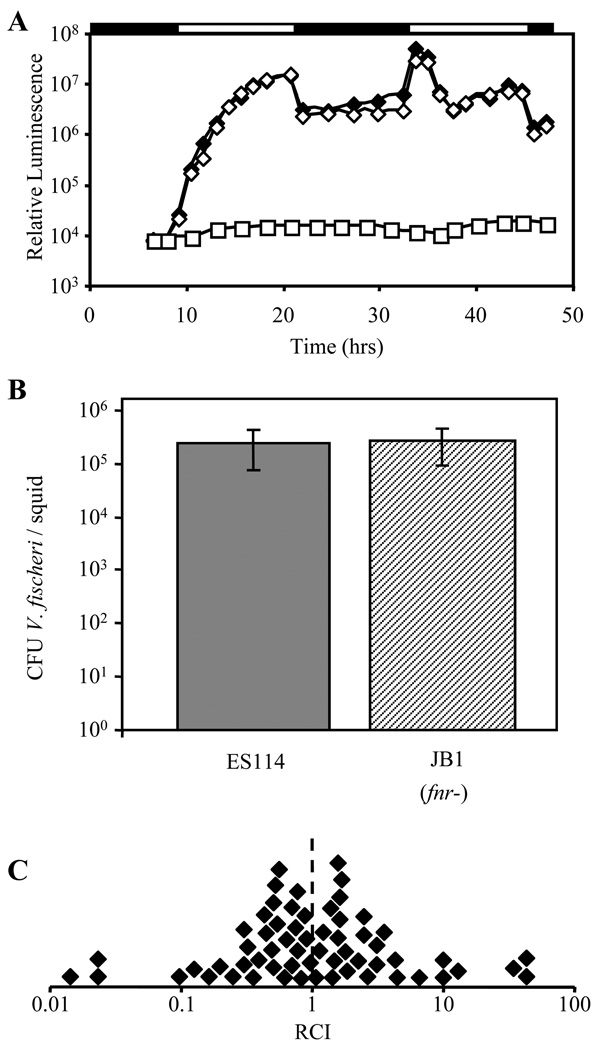
Vibrio fischeri induces both anaerobic respiration and bioluminescence during symbiotic infection. In many bacteria, the oxygen-sensitive regulator FNR activates anaerobic respiration, and a preliminary study using the light-generating lux genes from V. fischeri MJ1 cloned in Escherichia coli suggested that FNR stimulates bioluminescence. To test for FNR-mediated regulation of bioluminescence and anaerobic respiration in V. fischeri, we generated fnr mutants of V. fischeri strains MJ1 and ES114. In both strains, FNR was required for normal fumarate- and nitrate-dependent respiration. However, contrary to the report in transgenic E. coli, FNR mediated the repression of lux. ArcA represses bioluminescence, and P(arcA)-lacZ reporters showed reduced expression in fnr mutants, suggesting a possible indirect effect of FNR on bioluminescence via arcA. Finally, the fnr mutant of ES114 was not impaired in colonization of its host squid, Euprymna scolopes. This study extends the characterization of FNR to the Vibrionaceae and underscores the importance of studying lux regulation in its native background.
Figures
Similar articles
-
Effects of luxCDABEG induction in Vibrio fischeri: enhancement of symbiotic colonization and conditional attenuation of growth in culture.Arch Microbiol. 2008 Aug;190(2):169-83. doi: 10.1007/s00203-008-0387-1. Epub 2008 Jun 3. Arch Microbiol. 2008. PMID: 18521572 Free PMC article.
-
Bioluminescence in Vibrio fischeri is controlled by the redox-responsive regulator ArcA.Mol Microbiol. 2007 Jul;65(2):538-53. doi: 10.1111/j.1365-2958.2007.05809.x. Epub 2007 Jun 21. Mol Microbiol. 2007. PMID: 17590235
-
HbtR, a Heterofunctional Homolog of the Virulence Regulator TcpP, Facilitates the Transition between Symbiotic and Planktonic Lifestyles in Vibrio fischeri.mBio. 2020 Sep 1;11(5):e01624-20. doi: 10.1128/mBio.01624-20. mBio. 2020. PMID: 32873761 Free PMC article.
-
Quorum sensing in the squid-Vibrio symbiosis.Int J Mol Sci. 2013 Aug 7;14(8):16386-401. doi: 10.3390/ijms140816386. Int J Mol Sci. 2013. PMID: 23965960 Free PMC article. Review.
-
Vibrio fischeri: a model for host-associated biofilm formation.J Bacteriol. 2024 Feb 22;206(2):e0037023. doi: 10.1128/jb.00370-23. Epub 2024 Jan 25. J Bacteriol. 2024. PMID: 38270381 Free PMC article. Review.
Cited by
-
The crystal structure of the global anaerobic transcriptional regulator FNR explains its extremely fine-tuned monomer-dimer equilibrium.Sci Adv. 2015 Dec 4;1(11):e1501086. doi: 10.1126/sciadv.1501086. eCollection 2015 Dec. Sci Adv. 2015. PMID: 26665177 Free PMC article.
-
Contribution of rapid evolution of the luxR-luxI intergenic region to the diverse bioluminescence outputs of Vibrio fischeri strains isolated from different environments.Appl Environ Microbiol. 2011 Apr;77(7):2445-57. doi: 10.1128/AEM.02643-10. Epub 2011 Feb 11. Appl Environ Microbiol. 2011. PMID: 21317265 Free PMC article.
-
Rethinking the roles of CRP, cAMP, and sugar-mediated global regulation in the Vibrionaceae.Curr Genet. 2016 Feb;62(1):39-45. doi: 10.1007/s00294-015-0508-8. Epub 2015 Jul 28. Curr Genet. 2016. PMID: 26215147 Review.
-
Genomic Insights into Selenate Reduction by Anaerobacillus Species.Microorganisms. 2025 Mar 14;13(3):659. doi: 10.3390/microorganisms13030659. Microorganisms. 2025. PMID: 40142551 Free PMC article.
-
The iron-dependent regulator fur controls pheromone signaling systems and luminescence in the squid symbiont Vibrio fischeri ES114.Appl Environ Microbiol. 2013 Mar;79(6):1826-34. doi: 10.1128/AEM.03079-12. Epub 2013 Jan 11. Appl Environ Microbiol. 2013. PMID: 23315731 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
