

The human Mediator complex: a versatile, genome-wide regulator of transcription
- PMID: 20299225
- PMCID: PMC2891401
- DOI: 10.1016/j.tibs.2010.02.004
The human Mediator complex: a versatile, genome-wide regulator of transcription
Abstract
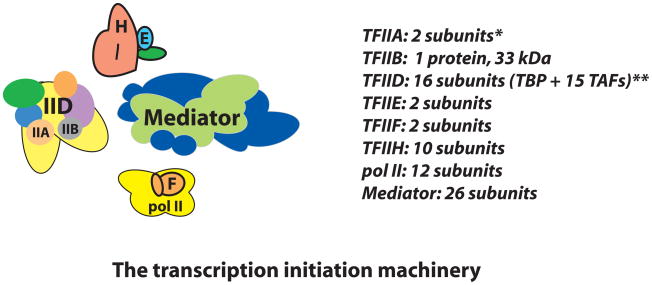
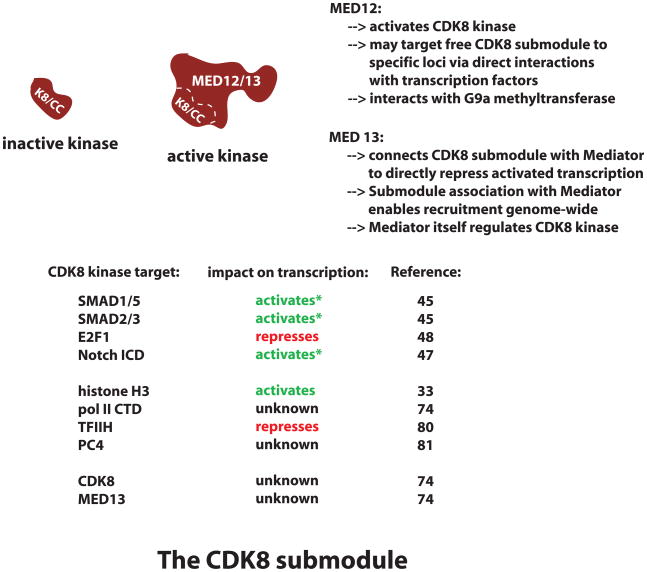
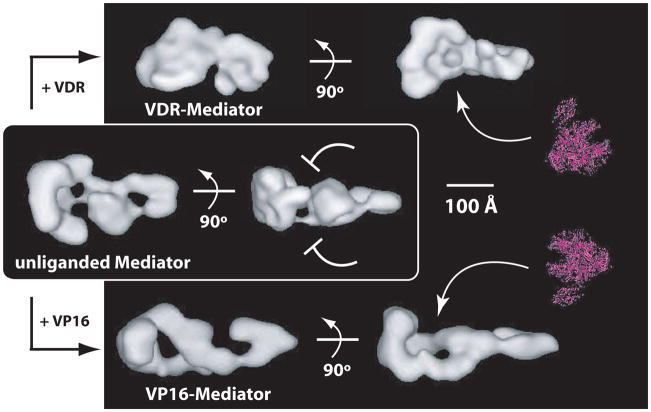
The Mediator complex interacts extensively with the RNA polymerase II enzyme and regulates its ability to express protein-coding genes. The mechanisms by which Mediator regulates gene expression remain poorly understood, in part because the structure of Mediator and even its composition can change, depending upon the promoter context. Combined with the sheer size of the human Mediator complex (26 subunits, 1.2 MDa), this structural adaptability bestows seemingly unlimited regulatory potential within the complex. Recent efforts to understand Mediator structure and function have identified expanded roles that include control of both pre- and post-initiation events; it is also evident that Mediator performs both general and gene-specific roles to regulate gene expression.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures


References
-
- Thompson CM, et al. A multisubunit complex associated with the RNA polymerase II CTD and TATA-binding protein in yeast. Cell. 1993;73:1361–1375. - PubMed
-
- Kim Y, et al. A multiprotein mediator of transcriptional activation and its interaction with the C-terminal repeat domain of RNA polymerase II. Cell. 1994;77:599–608. - PubMed
-
- Levine M, Tjian R. Transcription regulation and animal diversity. Nature. 2003;424:147–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
