

Retinal very long-chain PUFAs: new insights from studies on ELOVL4 protein
- PMID: 20299492
- PMCID: PMC2882734
- DOI: 10.1194/jlr.R005025
Retinal very long-chain PUFAs: new insights from studies on ELOVL4 protein
Abstract
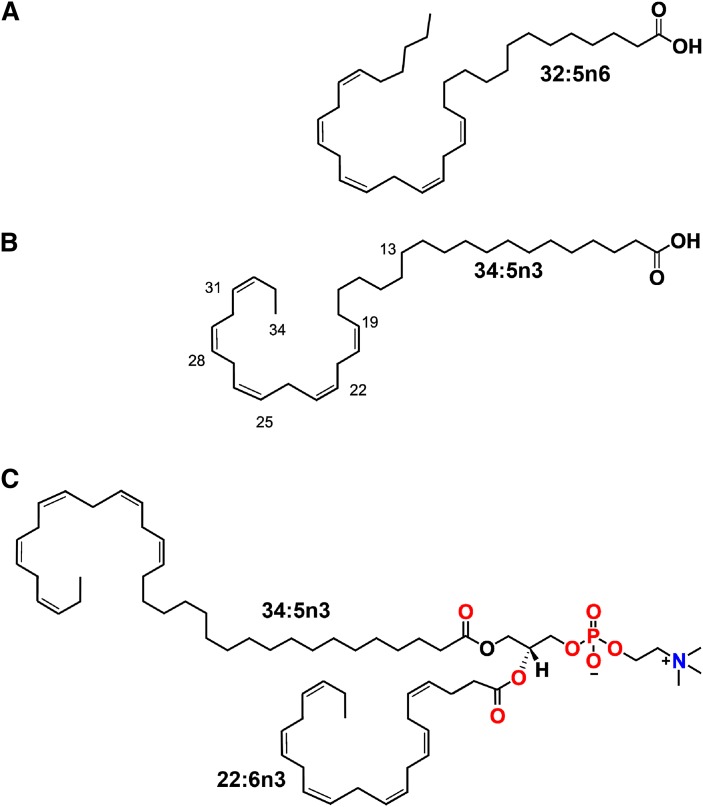
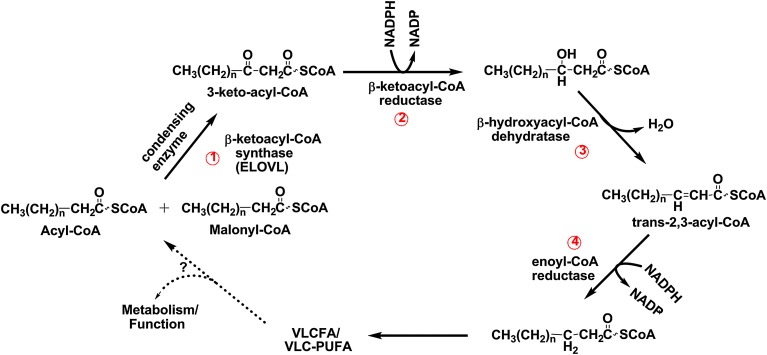
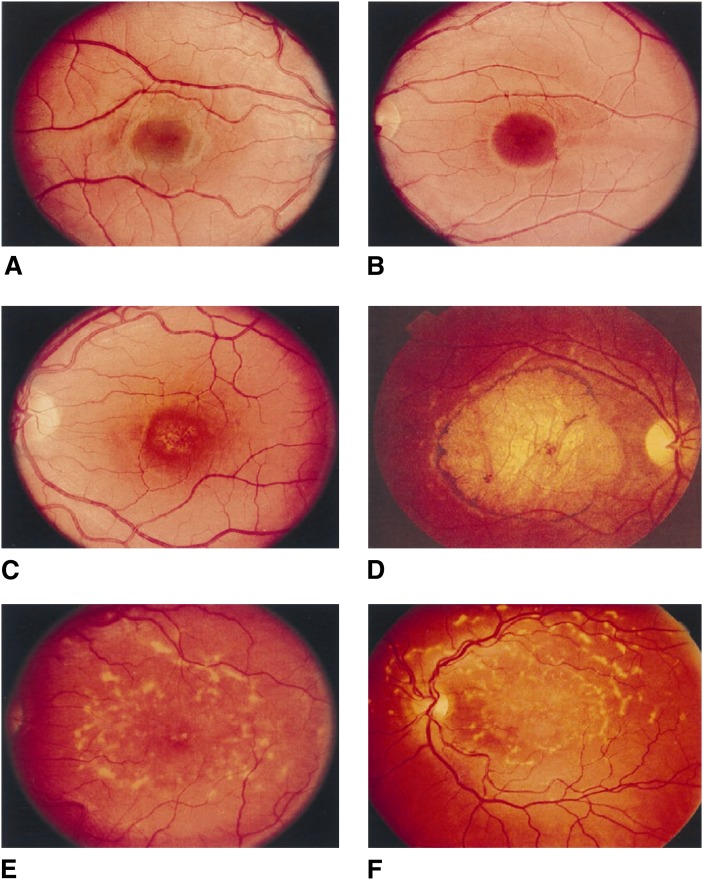
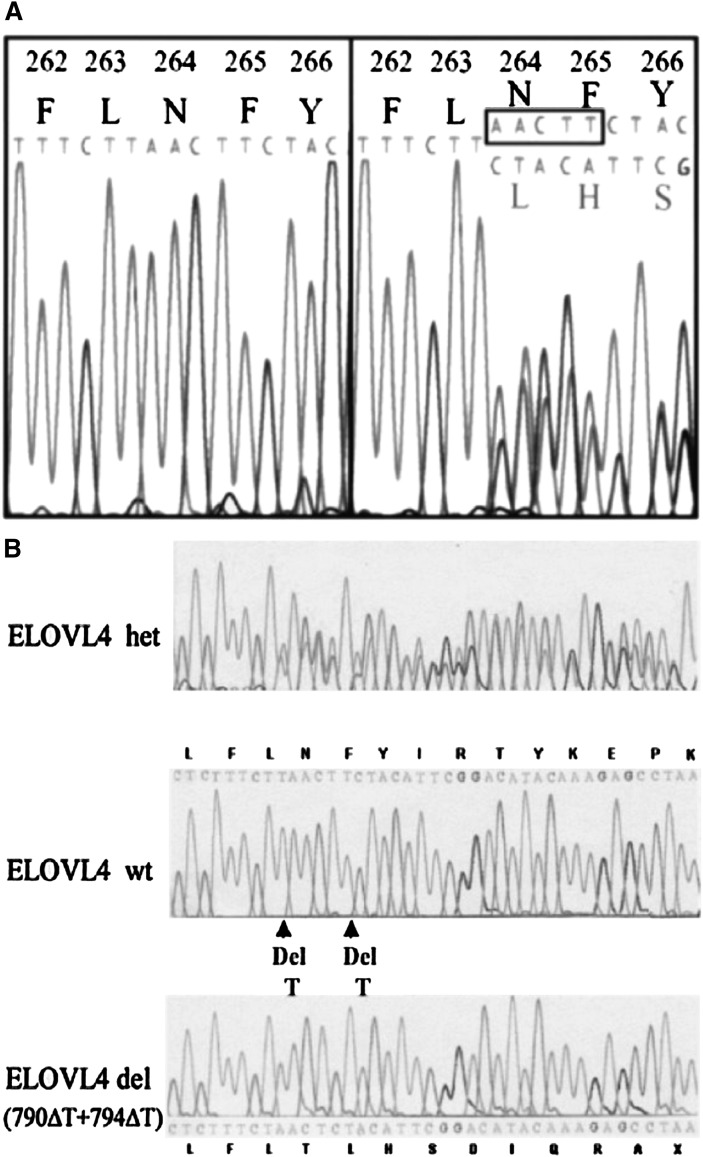
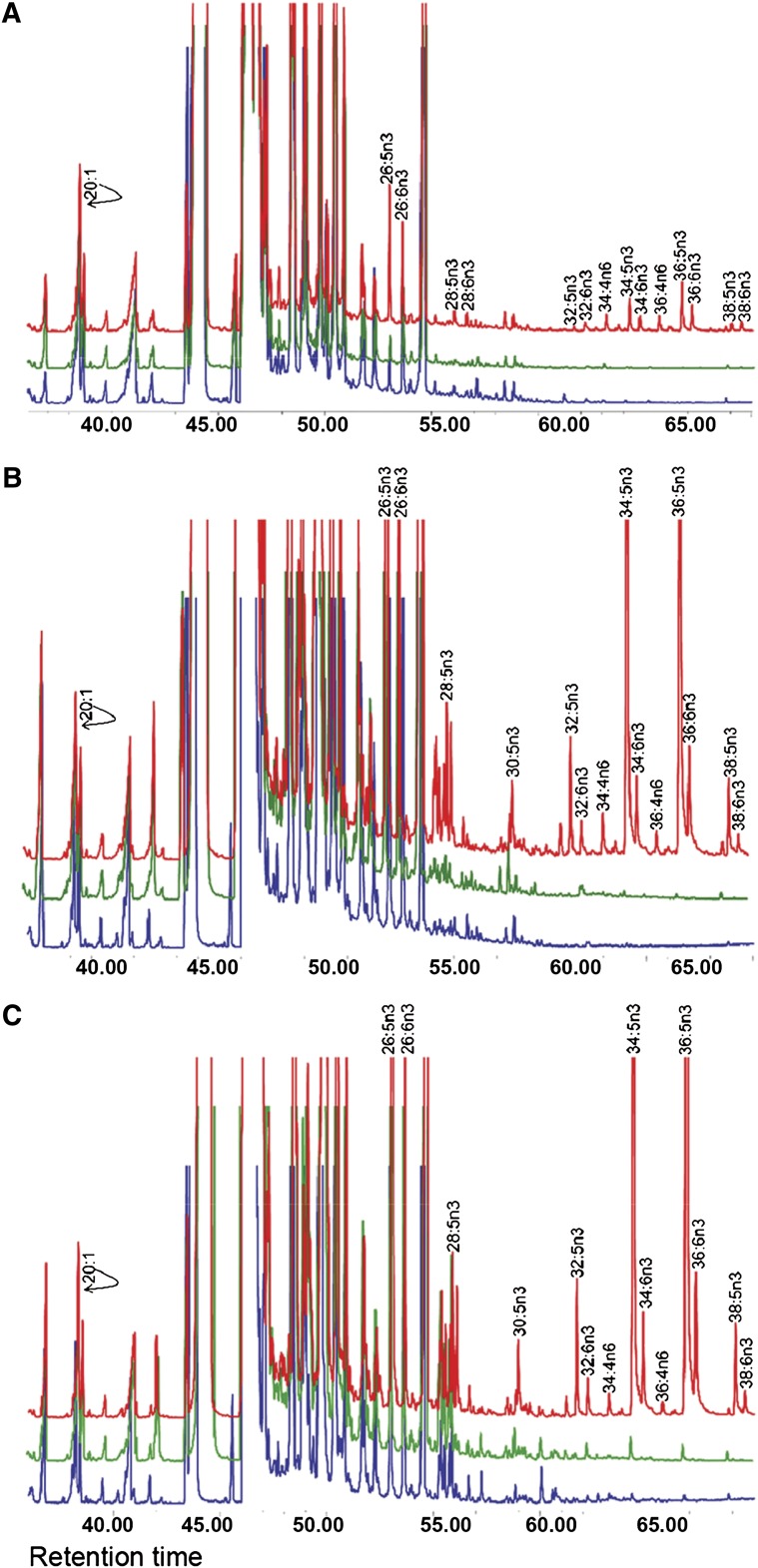
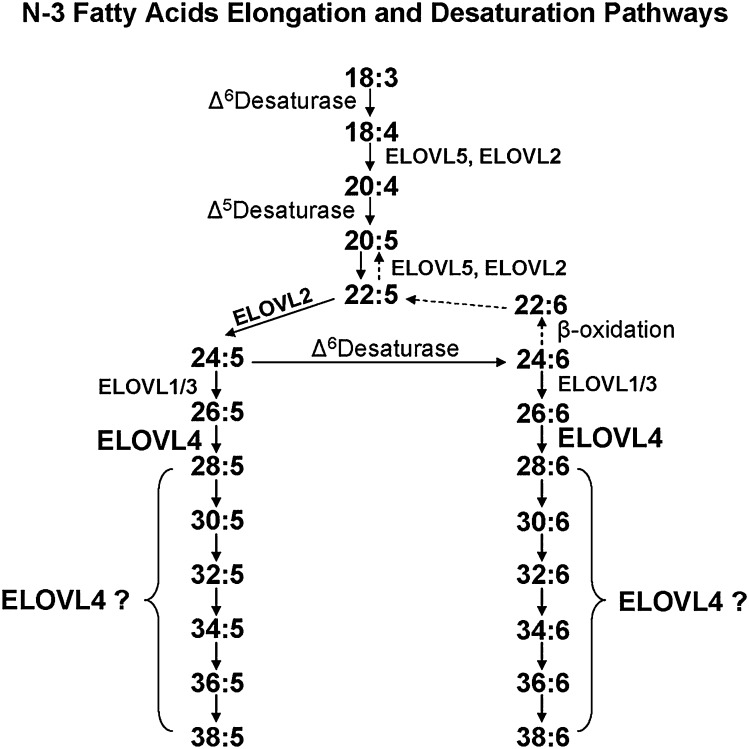
Compared with other mammalian tissues, retina is highly enriched in PUFA. Long-chain PUFA (LC-PUFA; C18-C24) are essential FAs that are enriched in the retina and are necessary for maintenance of normal retinal development and function. The retina, brain, and sperm also contain very LC-PUFA (VLC-PUFA; >C24). Although VLC-PUFA were discovered more than two decades ago, very little is known about their biosynthesis and functional roles in the retina. This is due mainly to intrinsic difficulties associated with working on these unusually long polyunsaturated hydrocarbon chains and their existence in small amounts. Recent studies on the FA elongase elongation of very long chain fatty acids-4 (ELOVL4) protein, however, suggest that VLC-PUFA probably play some uniquely important roles in the retina as well as the other tissues. Mutations in the ELOVL4 gene are found in patients with autosomal dominant Stargardt disease. Here, we review the recent literature on VLC-PUFA with special emphasis on the elongases responsible for their synthesis. We focus on a novel elongase, ELOVL4, involved in the synthesis of VLC-PUFA, and the importance of these FAs in maintaining the structural and functional integrity of retinal photoreceptors.
Figures
References
-
- Rezanka T. 1989. Very-long-chain fatty acids from the animal and plant kingdoms. Prog. Lipid Res. 28: 147–187. - PubMed
-
- Tvrzicka E., Rezanka T., Krijt J., Janousek V. 1988. Identification of very-long- chain fatty acids in rat and mouse harderian gland lipids by capillary gas chromatography-mass spectrometry. J. Chromatogr. 431: 231–238. - PubMed
-
- Poulos A. 1995. Very long chain fatty acids in higher animals–a review. Lipids. 30: 1–14. - PubMed
-
- Aveldano M. I. 1987. A novel group of very long chain polyenoic fatty acids in dipolyunsaturated phosphatidylcholines from vertebrate retina. J. Biol. Chem. 262: 1172–1179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
