

Tgf-Beta isoform specific regulation of airway inflammation and remodelling in a murine model of asthma
- PMID: 20300191
- PMCID: PMC2837347
- DOI: 10.1371/journal.pone.0009674
Tgf-Beta isoform specific regulation of airway inflammation and remodelling in a murine model of asthma
Abstract
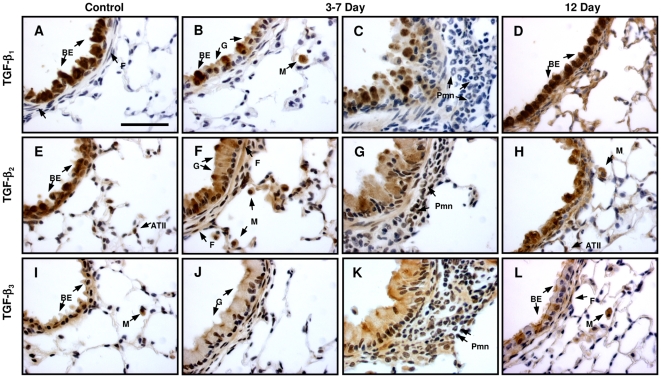
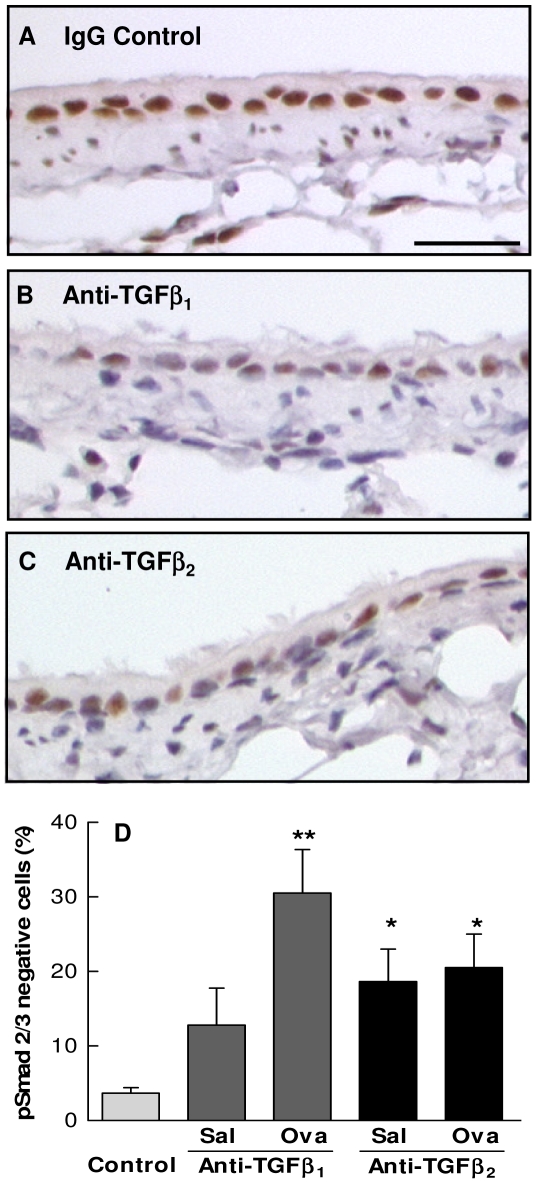
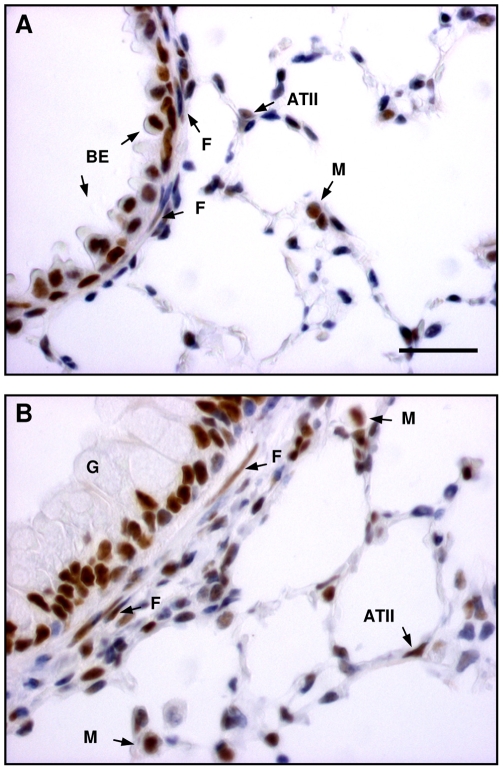
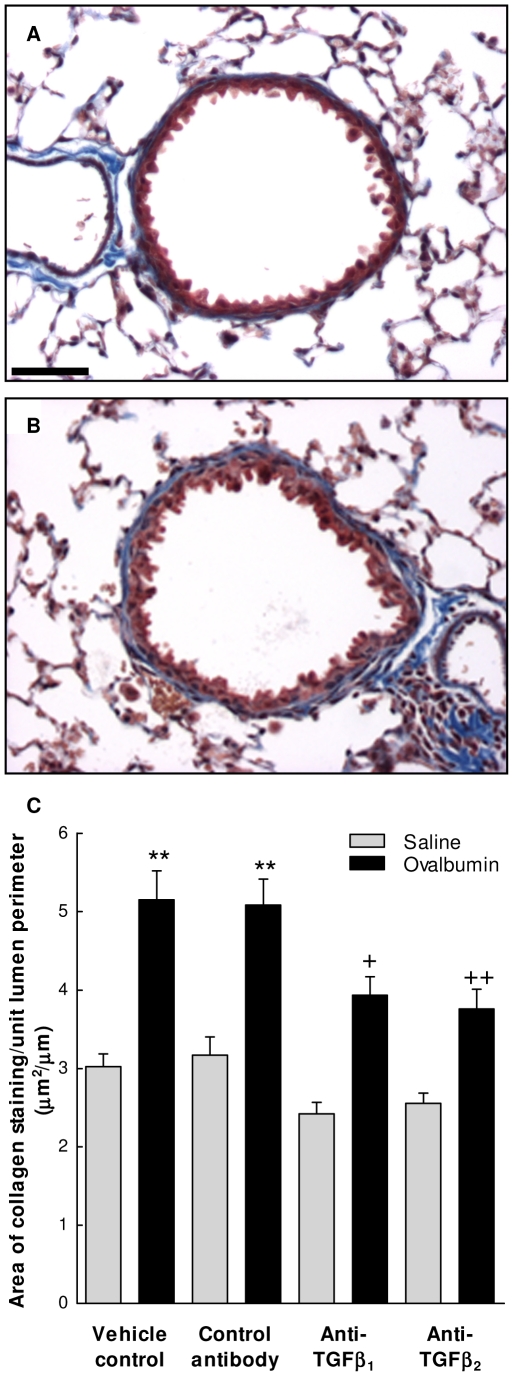
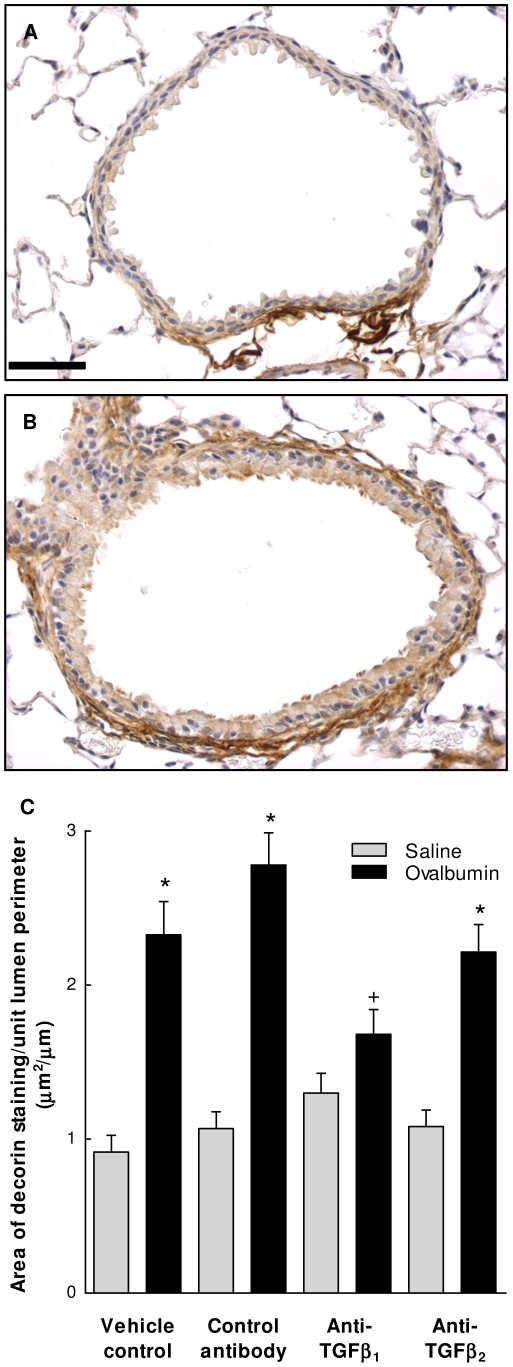
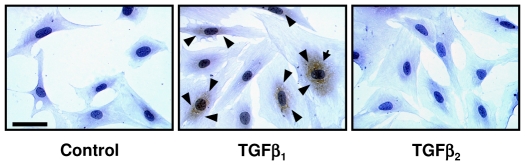
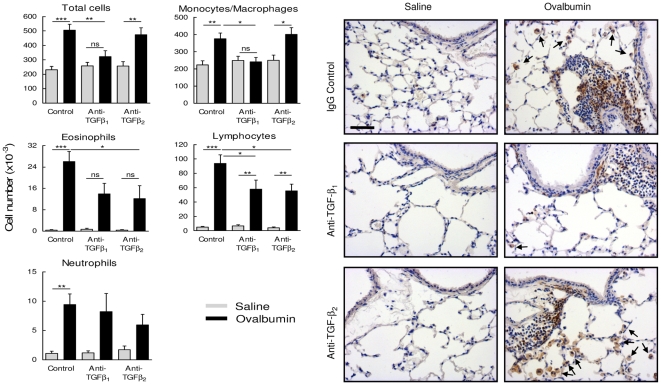
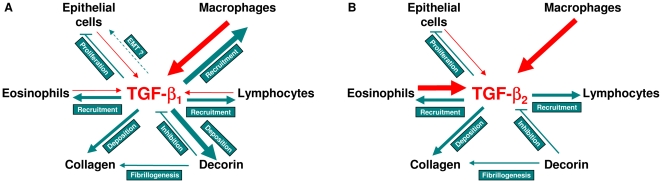
The TGF-beta family of mediators are thought to play important roles in the regulation of inflammation and airway remodelling in asthma. All three mammalian isoforms of TGF-beta, TGF-beta(1-3), are expressed in the airways and TGF-beta(1) and -beta(2) are increased in asthma. However, there is little information on the specific roles of individual TGF-beta isoforms. In this study we assess the roles of TGF-beta(1) and TGF-beta(2) in the regulation of allergen-induced airway inflammation and remodelling associated with asthma, using a validated murine model of ovalbumin sensitization and challenge, and isoform specific TGF-beta neutralising antibodies. Antibodies to both isoforms inhibited TGF-beta mediated Smad signalling. Anti-TGF-beta(1) and anti-TGF-beta(2) inhibited ovalbumin-induced sub-epithelial collagen deposition but anti-TGF-beta(1) also specifically regulated airway and fibroblast decorin deposition by TGF-beta(1). Neither antibody affected the allergen-induced increase in sub-epithelial fibroblast-like cells. Anti- TGF-beta(1) also specifically inhibited ovalbumin-induced increases in monocyte/macrophage recruitment. Whereas, both TGF-beta(1) and TGF-beta(2) were involved in regulating allergen-induced increases in eosinophil and lymphocyte numbers. These data show that TGF-beta(1) and TGF-beta(2) exhibit a combination of specific and shared roles in the regulation of allergen-induced airway inflammation and remodelling. They also provide evidence in support of the potential for therapeutic regulation of specific subsets of cells and extracellular matrix proteins associated with inflammation and remodelling in airway diseases such as asthma and COPD, as well as other fibroproliferative diseases.
Conflict of interest statement
Figures
Similar articles
-
Respiratory syncytial virus infection provokes airway remodelling in allergen-exposed mice in absence of prior allergen sensitization.Clin Exp Allergy. 2008 Jun;38(6):1016-24. doi: 10.1111/j.1365-2222.2008.02974.x. Clin Exp Allergy. 2008. PMID: 18498543 Free PMC article.
-
Manipulation of allergen-induced airway remodeling by treatment with anti-TGF-beta antibody: effect on the Smad signaling pathway.J Immunol. 2005 May 1;174(9):5774-80. doi: 10.4049/jimmunol.174.9.5774. J Immunol. 2005. PMID: 15843580
-
Expression and activation of TGF-beta isoforms in acute allergen-induced remodelling in asthma.Thorax. 2007 Apr;62(4):307-13. doi: 10.1136/thx.2006.063487. Epub 2007 Jan 24. Thorax. 2007. PMID: 17251317 Free PMC article.
-
TGF-beta: its role in asthma and therapeutic potential.Curr Drug Targets. 2006 May;7(5):547-65. doi: 10.2174/138945006776818692. Curr Drug Targets. 2006. PMID: 16719766 Review.
-
Interaction of tgf-beta with immune cells in airway disease.Curr Mol Med. 2008 Aug;8(5):427-36. doi: 10.2174/156652408785160943. Curr Mol Med. 2008. PMID: 18691070 Review.
Cited by
-
Transcriptome analysis of porcine M. semimembranosus divergent in intramuscular fat as a consequence of dietary protein restriction.BMC Genomics. 2013 Jul 6;14:453. doi: 10.1186/1471-2164-14-453. BMC Genomics. 2013. PMID: 23829541 Free PMC article.
-
Transforming growth factor-β-induced differentiation of airway smooth muscle cells is inhibited by fibroblast growth factor-2.Am J Respir Cell Mol Biol. 2013 Mar;48(3):346-53. doi: 10.1165/rcmb.2012-0151OC. Epub 2012 Dec 13. Am J Respir Cell Mol Biol. 2013. PMID: 23239497 Free PMC article.
-
Aerosolised Mesenchymal Stem Cells Expressing Angiopoietin-1 Enhances Airway Repair.Stem Cell Rev Rep. 2019 Feb;15(1):112-125. doi: 10.1007/s12015-018-9844-7. Stem Cell Rev Rep. 2019. PMID: 30178289
-
Vitamin D regulating TGF-β induced epithelial-mesenchymal transition.Respir Res. 2014 Nov 21;15(1):146. doi: 10.1186/s12931-014-0146-6. Respir Res. 2014. PMID: 25413472 Free PMC article.
-
Genotype-dependent effects of TGF-β1 on mast cell function: targeting the Stat5 pathway.J Immunol. 2013 Nov 1;191(9):4505-13. doi: 10.4049/jimmunol.1202723. Epub 2013 Sep 25. J Immunol. 2013. PMID: 24068671 Free PMC article.
References
-
- Howell JE, McAnulty RJ. TGF-beta: its role in asthma and therapeutic potential. Curr Drug Targets. 2006;7:547–565. - PubMed
-
- Roche WR, Beasley R, Williams JH, Holgate ST. Subepithelial fibrosis in the bronchi of asthmatics. Lancet. 1989;1:520–524. - PubMed
-
- Roberts CR. Is asthma a fibrotic disease? Chest. 1995;107:111S–117S. - PubMed
-
- Hoshino M, Nakamura Y, Sim J, Shimojo J, Isogai S. Bronchial subepithelial fibrosis and expression of matrix metalloproteinase-9 in asthmatic airway inflammation. J Allergy Clin Immunol. 1998;102:783–788. - PubMed
-
- Huang J, Olivenstein R, Taha R, Hamid Q, Ludwig M. Enhanced proteoglycan deposition in the airway wall of atopic asthmatics. Am J Respir Crit Care Med. 1999;160:725–729. - PubMed
