

Regenerative medicine for retinal diseases: activating endogenous repair mechanisms
- PMID: 20303826
- PMCID: PMC2854262
- DOI: 10.1016/j.molmed.2010.02.003
Regenerative medicine for retinal diseases: activating endogenous repair mechanisms
Abstract
The retina is subject to degenerative diseases that often lead to significant visual impairment. Non-mammalian vertebrates have the remarkable ability to replace neurons lost through damage. Fish, and to a limited extent birds, replace lost neurons by the dedifferentiation of Müller glia to a progenitor state followed by the replication of these neuronal progenitor cells. Over the past five years, studies have investigated whether regeneration can be stimulated in the mouse and rat retina. Several groups have reported that at least some types of neurons can be regenerated in the mammalian retina in vivo or in vitro, and that the regeneration of neurons can be stimulated using growth factors, transcription factors or subtoxic levels of excitatory amino acids. These recent results suggest that some part of the regenerative program that occurs in non-mammalian vertebrates remains in the mammalian retina, and could provide a basis to develop new strategies for retinal repair in patients with retinal degenerations.
Figures
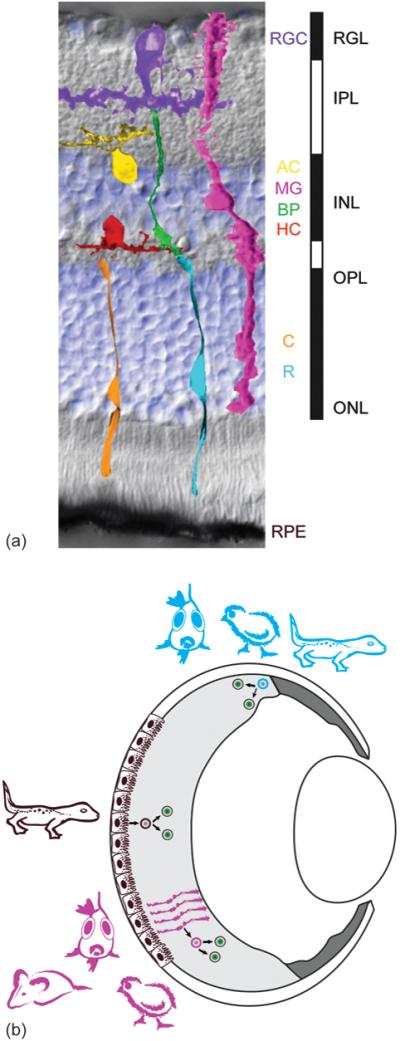
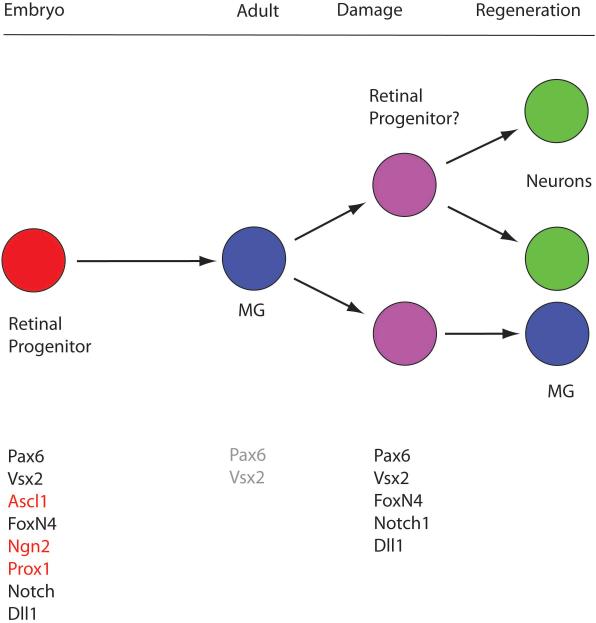
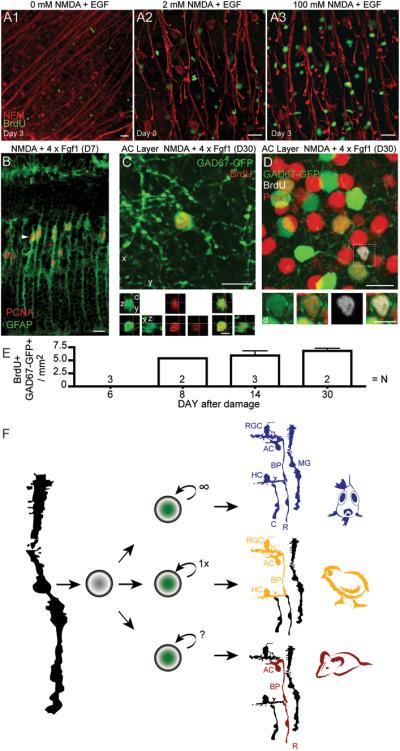
References
-
- Jones BW, et al. Retinal remodeling triggered by photoreceptor degenerations. J Comp Neurol. 2003;464:1–16. - PubMed
-
- Fariss RN, et al. Abnormalities in rod photoreceptors, amacrine cells, and horizontal cells in human retinas with retinitis pigmentosa. Am J Ophthalmol. 2000;129:215–223. - PubMed
-
- Moshiri A, et al. Retinal stem cells and regeneration. Int J Dev Biol. 2004;48:1003–1014. - PubMed
-
- Tropepe V, et al. Retinal stem cells in the adult mammalian eye. Science. 2000;287:2032–2036. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
