

Interaction of 18-methoxycoronaridine with nicotinic acetylcholine receptors in different conformational states
- PMID: 20303928
- PMCID: PMC3787694
- DOI: 10.1016/j.bbamem.2010.03.013
Interaction of 18-methoxycoronaridine with nicotinic acetylcholine receptors in different conformational states
Abstract
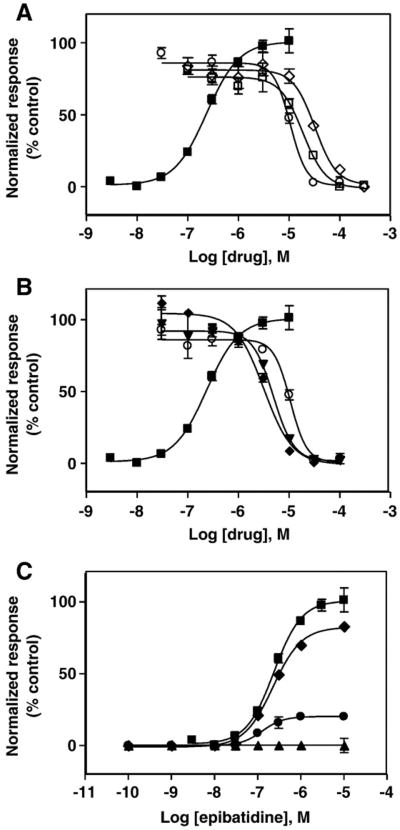
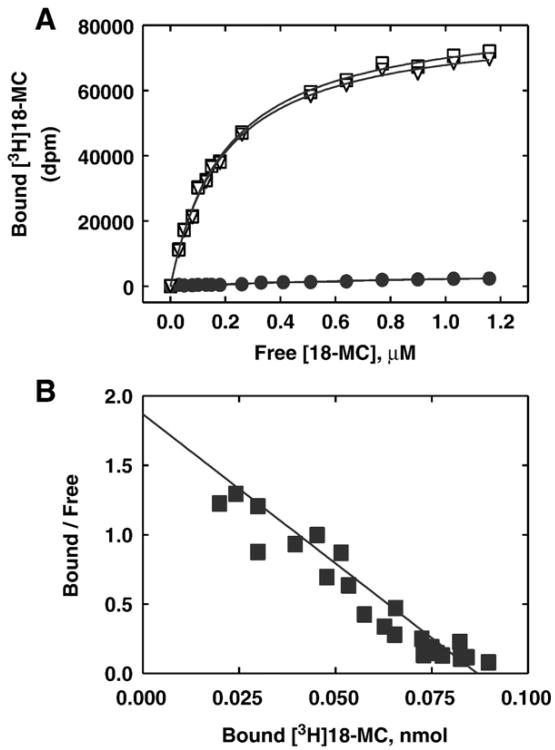
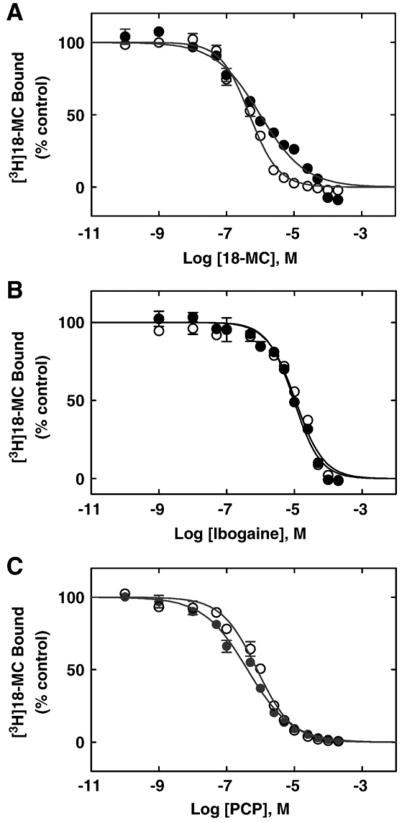
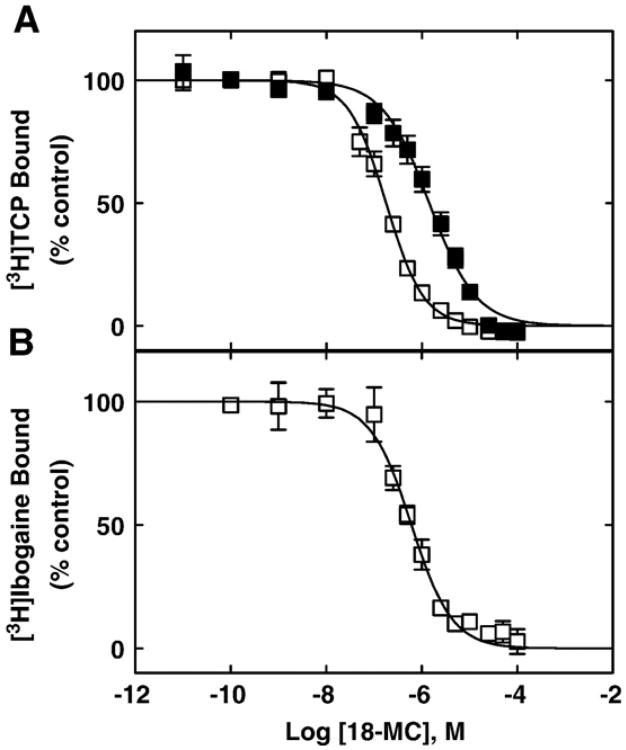
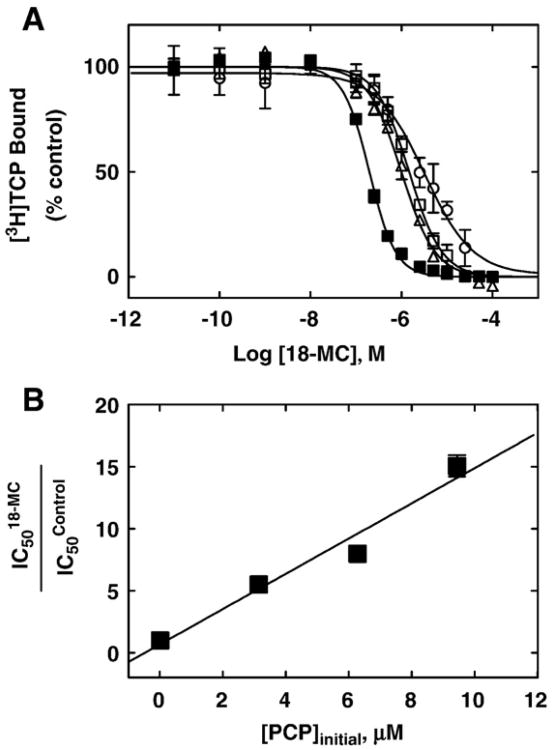
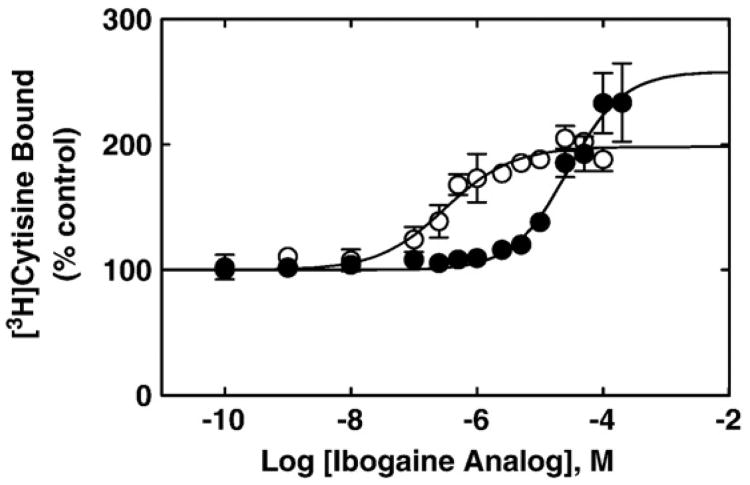
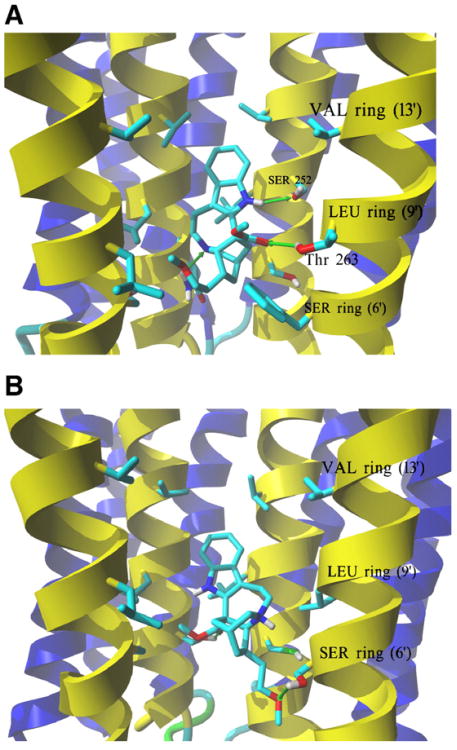
The interaction of 18-methoxycoronaridine (18-MC) with nicotinic acetylcholine receptors (AChRs) was compared with that for ibogaine and phencyclidine (PCP). The results established that 18-MC: (a) is more potent than ibogaine and PCP inhibiting (+/-)-epibatidine-induced AChR Ca(2+) influx. The potency of 18-MC is increased after longer pre-incubation periods, which is in agreement with the enhancement of [(3)H]cytisine binding to resting but activatable Torpedo AChRs, (b) binds to a single site in the Torpedo AChR with high affinity and inhibits [(3)H]TCP binding to desensitized AChRs in a steric fashion, suggesting the existence of overlapping sites. This is supported by our docking results indicating that 18-MC interacts with a domain located between the serine (position 6') and valine (position 13') rings, and (c) inhibits [(3)H]TCP, [(3)H]ibogaine, and [(3)H]18-MC binding to desensitized AChRs with higher affinity compared to resting AChRs. This can be partially attributed to a slower dissociation rate from the desensitized AChR compared to that from the resting AChR. The enthalpic contribution is more important than the entropic contribution when 18-MC binds to the desensitized AChR compared to that for the resting AChR, and vice versa. Ibogaine analogs inhibit the AChR by interacting with a luminal domain that is shared with PCP, and by inducing desensitization.
Figures
References
-
- Arias HR. Thermodynamics of nicotinic receptor interactions. In: Raffa RB, editor. Drug-Receptor Thermodynamics: Introduction and Applications. John Wiley & Sons, Ltd; USA: 2001. pp. 293–358.
-
- Arias HR. Ligand-gated ion channel receptor superfamilies. In: Arias HR, editor. Biological and Biophysical Aspects of Ligand-Gated Ion Channel Receptor Superfamilies. Research Signpost; Kerala, India: 2006. pp. 1–25.
-
- Arias HR, Bhumireddy P, Bouzat C. Molecular mechanisms and binding site locations for noncompetitive antagonists of nicotinic acetylcholine receptors. Int J Biochem Cell Biol. 2006;38:1254–1276. - PubMed
-
- Sine SM, Engel AG. Recent advances in Cys-loop receptor structure and function. Nature. 2006;440:448–455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
