

A mobile threat to genome stability: The impact of non-LTR retrotransposons upon the human genome
- PMID: 20307669
- PMCID: PMC2925057
- DOI: 10.1016/j.semcancer.2010.03.001
A mobile threat to genome stability: The impact of non-LTR retrotransposons upon the human genome
Abstract
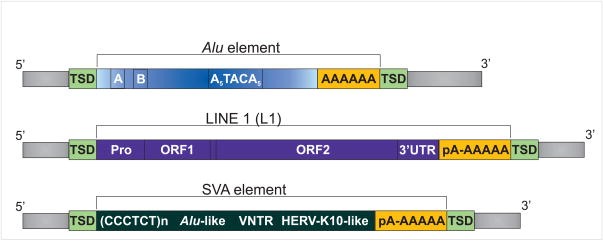
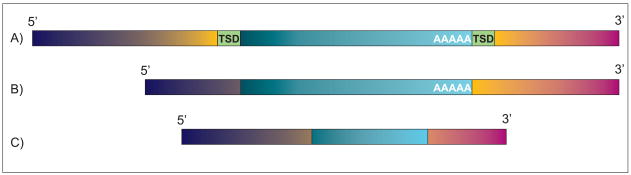
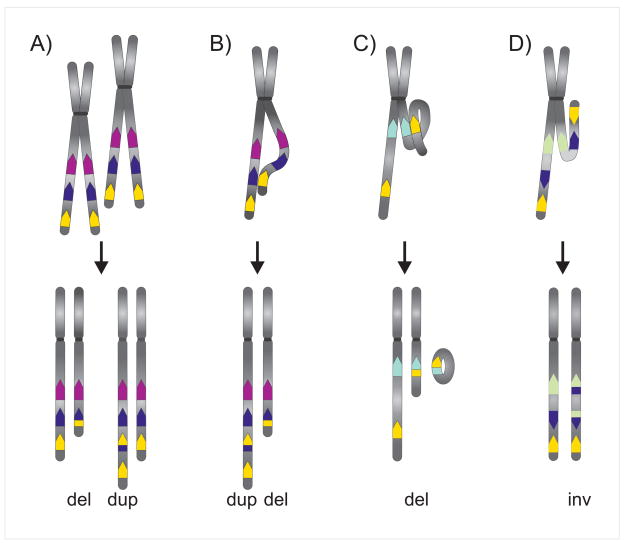
It is now commonly agreed that the human genome is not the stable entity originally presumed. Deletions, duplications, inversions, and insertions are common, and contribute significantly to genomic structural variations (SVs). Their collective impact generates much of the inter-individual genomic diversity observed among humans. Not only do these variations change the structure of the genome; they may also have functional implications, e.g. altered gene expression. Some SVs have been identified as the cause of genetic disorders, including cancer predisposition. Cancer cells are notorious for their genomic instability, and often show genomic rearrangements at the microscopic and submicroscopic level to which transposable elements (TEs) contribute. Here, we review the role of TEs in genome instability, with particular focus on non-LTR retrotransposons. Currently, three non-LTR retrotransposon families - long interspersed element 1 (L1), SVA (short interspersed element (SINE-R), variable number of tandem repeats (VNTR), and Alu), and Alu (a SINE) elements - mobilize in the human genome, and cause genomic instability through both insertion- and post-insertion-based mutagenesis. Due to the abundance and high sequence identity of TEs, they frequently mislead the homologous recombination repair pathway into non-allelic homologous recombination, causing deletions, duplications, and inversions. While less comprehensively studied, non-LTR retrotransposon insertions and TE-mediated rearrangements are probably more common in cancer cells than in healthy tissue. This may be at least partially attributed to the commonly seen global hypomethylation as well as general epigenetic dysfunction of cancer cells. Where possible, we provide examples that impact cancer predisposition and/or development.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare that there are no conflicts of interest.
Figures
Similar articles
-
All y'all need to know 'bout retroelements in cancer.Semin Cancer Biol. 2010 Aug;20(4):200-10. doi: 10.1016/j.semcancer.2010.06.001. Epub 2010 Jun 25. Semin Cancer Biol. 2010. PMID: 20600922 Free PMC article. Review.
-
Exceptional diversity, non-random distribution, and rapid evolution of retroelements in the B73 maize genome.PLoS Genet. 2009 Nov;5(11):e1000732. doi: 10.1371/journal.pgen.1000732. Epub 2009 Nov 20. PLoS Genet. 2009. PMID: 19936065 Free PMC article.
-
Do genetic recombination and gene density shape the pattern of DNA elimination in rice long terminal repeat retrotransposons?Genome Res. 2009 Dec;19(12):2221-30. doi: 10.1101/gr.083899.108. Epub 2009 Sep 29. Genome Res. 2009. PMID: 19789376 Free PMC article.
-
Constant conflict between Gypsy LTR retrotransposons and CHH methylation within a stress-adapted mangrove genome.New Phytol. 2018 Nov;220(3):922-935. doi: 10.1111/nph.15209. Epub 2018 May 15. New Phytol. 2018. PMID: 29762876
-
Inviting instability: Transposable elements, double-strand breaks, and the maintenance of genome integrity.Mutat Res. 2007 Mar 1;616(1-2):46-59. doi: 10.1016/j.mrfmmm.2006.11.021. Epub 2006 Dec 8. Mutat Res. 2007. PMID: 17157332 Free PMC article. Review.
Cited by
-
Detailed Analysis of Focal Chromosome Arm 1q and 6p Amplifications in Urothelial Carcinoma Reveals Complex Genomic Events on 1q, and SOX4 as a Possible Auxiliary Target on 6p.PLoS One. 2013 Jun 18;8(6):e67222. doi: 10.1371/journal.pone.0067222. Print 2013. PLoS One. 2013. PMID: 23825644 Free PMC article.
-
Long interspersed element-1 is differentially regulated by food-borne carcinogens via the aryl hydrocarbon receptor.Oncogene. 2013 Oct 10;32(41):4903-12. doi: 10.1038/onc.2012.516. Epub 2012 Dec 3. Oncogene. 2013. PMID: 23208499 Free PMC article.
-
Roles of Human Endogenous Retroviruses and Endogenous Virus-Like Elements in Cancer Development and Innate Immunity.Biomolecules. 2023 Nov 24;13(12):1706. doi: 10.3390/biom13121706. Biomolecules. 2023. PMID: 38136578 Free PMC article. Review.
-
Endogenous Retroelements and the Host Innate Immune Sensors.Adv Immunol. 2016;132:47-69. doi: 10.1016/bs.ai.2016.07.001. Epub 2016 Aug 23. Adv Immunol. 2016. PMID: 27769507 Free PMC article. Review.
-
Massive gene amplification on a recently formed Drosophila Y chromosome.Nat Ecol Evol. 2019 Nov;3(11):1587-1597. doi: 10.1038/s41559-019-1009-9. Epub 2019 Oct 28. Nat Ecol Evol. 2019. PMID: 31666742 Free PMC article.
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, et al. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921. - PubMed
-
- Sebat J, Lakshmi B, Troge J, Alexander J, Young J, Lundin P, et al. Large-scale copy number polymorphism in the human genome. Science. 2004;305:525–8. - PubMed
-
- Iafrate AJ, Feuk L, Rivera MN, Listewnik ML, Donahoe PK, Qi Y, et al. Detection of large-scale variation in the human genome. Nat Genet. 2004;36:949–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
