

Temporin-SHf, a new type of phe-rich and hydrophobic ultrashort antimicrobial peptide
- PMID: 20308076
- PMCID: PMC2878016
- DOI: 10.1074/jbc.M109.097204
Temporin-SHf, a new type of phe-rich and hydrophobic ultrashort antimicrobial peptide
Abstract
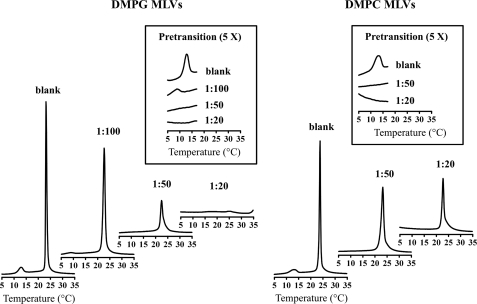
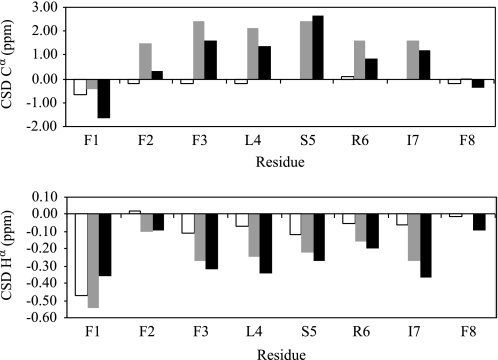
Because issues of cost and bioavailability have hampered the development of gene-encoded antimicrobial peptides to combat infectious diseases, short linear peptides with high microbial cell selectivity have been recently considered as antibiotic substitutes. A new type of short antimicrobial peptide, designated temporin-SHf, was isolated and cloned from the skin of the frog Pelophylax saharica. Temporin-SHf has a highly hydrophobic sequence (FFFLSRIFa) and possesses the highest percentage of Phe residues of any known peptide or protein. Moreover, it is the smallest natural linear antimicrobial peptide found to date, with only eight residues. Despite its small size and hydrophobicity, temporin-SHf has broad-spectrum microbicidal activity against Gram-positive and Gram-negative bacteria and yeasts, with no hemolytic activity. CD and NMR spectroscopy combined with restrained molecular dynamics calculations showed that the peptide adopts a well defined non-amphipathic alpha-helical structure from residue 3 to 8, when bound to zwitterionic dodecyl phosphocholine or anionic SDS micelles. Relaxation enhancement caused by paramagnetic probes showed that the peptide adopts nearly parallel orientations to the micelle surface and that the helical structure is stabilized by a compact hydrophobic core on one face that penetrates into the micelle interior. Differential scanning calorimetry on multilamellar vesicles combined with membrane permeabilization assays on bacterial cells indicated that temporin-SHf disrupts the acyl chain packing of anionic lipid bilayers, thereby triggering local cracks and microbial membrane disintegration through a detergent-like effect probably via the carpet mechanism. The short length, compositional simplicity, and broad-spectrum activity of temporin-SHf make it an attractive candidate to develop new antibiotic agents.
Figures








Similar articles
-
Solution structure and model membrane interactions of temporins-SH, antimicrobial peptides from amphibian skin. A NMR spectroscopy and differential scanning calorimetry study.Biochemistry. 2008 Oct 7;47(40):10513-25. doi: 10.1021/bi8006884. Epub 2008 Sep 17. Biochemistry. 2008. PMID: 18795798
-
Disulfide engineering on temporin-SHf: Stabilizing the bioactive conformation of an ultra-short antimicrobial peptide.Chem Biol Drug Des. 2019 Sep;94(3):1634-1646. doi: 10.1111/cbdd.13525. Epub 2019 Aug 4. Chem Biol Drug Des. 2019. PMID: 30924306
-
Structure-Activity Relationship-based Optimization of Small Temporin-SHf Analogs with Potent Antibacterial Activity.ACS Chem Biol. 2015 Oct 16;10(10):2257-66. doi: 10.1021/acschembio.5b00495. Epub 2015 Jul 30. ACS Chem Biol. 2015. PMID: 26181487
-
Solution NMR studies of amphibian antimicrobial peptides: linking structure to function?Biochim Biophys Acta. 2009 Aug;1788(8):1639-55. doi: 10.1016/j.bbamem.2009.01.002. Epub 2009 Jan 15. Biochim Biophys Acta. 2009. PMID: 19272309 Review.
-
Biological activity and structural aspects of PGLa interaction with membrane mimetic systems.Biochim Biophys Acta. 2009 Aug;1788(8):1656-66. doi: 10.1016/j.bbamem.2009.05.012. Epub 2009 May 29. Biochim Biophys Acta. 2009. PMID: 19481533 Review.
Cited by
-
Effect of Disulfide Cyclization of Ultrashort Cationic Lipopeptides on Antimicrobial Activity and Cytotoxicity.Int J Mol Sci. 2020 Sep 29;21(19):7208. doi: 10.3390/ijms21197208. Int J Mol Sci. 2020. PMID: 33003569 Free PMC article.
-
In vitro and intracellular activities of frog skin temporins against Legionella pneumophila and its eukaryotic hosts.Sci Rep. 2020 Mar 4;10(1):3978. doi: 10.1038/s41598-020-60829-2. Sci Rep. 2020. PMID: 32132569 Free PMC article.
-
Cobra cytotoxins: structural organization and antibacterial activity.Acta Naturae. 2014 Jul;6(3):11-8. Acta Naturae. 2014. PMID: 25349711 Free PMC article.
-
Temporins: Multifunctional Peptides from Frog Skin.Int J Mol Sci. 2023 Mar 12;24(6):5426. doi: 10.3390/ijms24065426. Int J Mol Sci. 2023. PMID: 36982501 Free PMC article. Review.
-
An active machine learning discovery platform for membrane-disrupting and pore-forming peptides.Phys Chem Chem Phys. 2024 Jun 26;26(25):17745-17752. doi: 10.1039/d4cp01404a. Phys Chem Chem Phys. 2024. PMID: 38873737 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
