

Estrogen receptor ESR1 controls cell migration by repressing chemokine receptor CXCR4 in the zebrafish posterior lateral line system
- PMID: 20308561
- PMCID: PMC2851976
- DOI: 10.1073/pnas.0909998107
Estrogen receptor ESR1 controls cell migration by repressing chemokine receptor CXCR4 in the zebrafish posterior lateral line system
Abstract
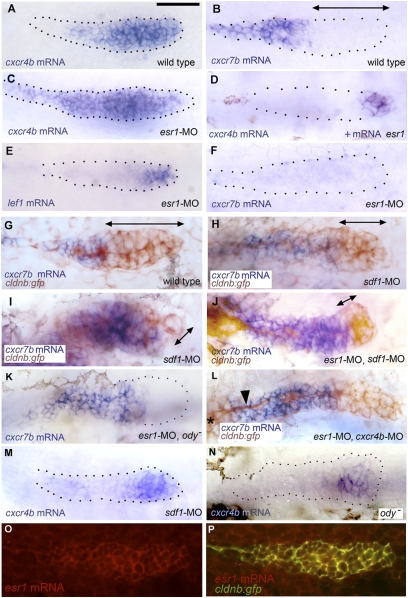
The primordium that generates the embryonic posterior lateral line of zebrafish migrates from the head to the tip of the tail along a trail of SDF1-producing cells. This migration critically depends on the presence of the SDF1 receptor CXCR4 in the leading region of the primordium and on the presence of a second SDF1 receptor, CXCR7, in the trailing region of the primordium. Here we show that inactivation of the estrogen receptor ESR1 results in ectopic expression of cxcr4b throughout the primordium, whereas ESR1 overexpression results in a reciprocal reduction in the domain of cxcr4b expression, suggesting that ESR1 acts as a repressor of cxcr4b. This finding could explain why estrogens significantly decrease the metastatic ability of ESR-positive breast cancer cells. ESR1 inactivation also leads to extinction of cxcr7b expression in the trailing cells of the migrating primordium; this effect is indirect, however, and due to the down-regulation of cxcr7b by ectopic SDF1/CXCR4 signaling in the trailing region. Both ESR1 inactivation and overexpression result in aborted migration, confirming the importance of this receptor in the control of SDF1-dependent migration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Ghysen A, Dambly-Chaudière C. The lateral line microcosmos. Genes Dev. 2007;21:2118–2130. - PubMed
-
- Metcalfe WK. Sensory neuron growth cones comigrate with posterior lateral line primordial cells in zebrafish. J Comp Neurol. 1985;238:218–224. - PubMed
-
- Gompel N, et al. Pattern formation in the lateral line of zebrafish. Mech Dev. 2001;105:69–77. - PubMed
-
- Li Q, Shirabe K, Kuwada JY. Chemokine signaling regulates sensory cell migration in zebrafish. Dev Biol. 2004;269:123–136. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
