

Imaging extrasynaptic glutamate dynamics in the brain
- PMID: 20308566
- PMCID: PMC2851965
- DOI: 10.1073/pnas.0913154107
Imaging extrasynaptic glutamate dynamics in the brain
Abstract
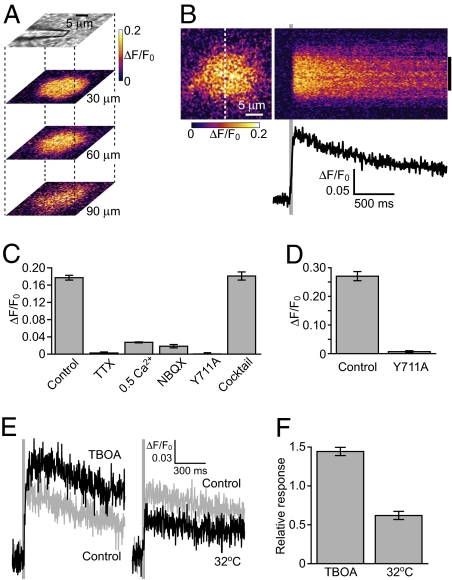
Glutamate is the major neurotransmitter in the brain, mediating point-to-point transmission across the synaptic cleft in excitatory synapses. Using a glutamate imaging method with fluorescent indicators, we show that synaptic activity generates extrasynaptic glutamate dynamics in the vicinity of active synapses. These glutamate dynamics had magnitudes and durations sufficient to activate extrasynaptic glutamate receptors in brain slices. We also observed crosstalk between synapses--i.e., summation of glutamate released from neighboring synapses. Furthermore, we successfully observed that sensory input from the extremities induced extrasynaptic glutamate dynamics within the appropriate sensory area of the cerebral cortex in vivo. Thus, the present study clarifies the spatiotemporal features of extrasynaptic glutamate dynamics, and opens up an avenue to directly visualizing synaptic activity in live animals.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Kullmann DM, Erdemli G, Asztély F. LTP of AMPA and NMDA receptor-mediated signals: evidence for presynaptic expression and extrasynaptic glutamate spill-over. Neuron. 1996;17:461–474. - PubMed
-
- Barbour B, Häusser M. Intersynaptic diffusion of neurotransmitter. Trends Neurosci. 1997;20:377–384. - PubMed
-
- Scanziani M, Salin PA, Vogt KE, Malenka RC, Nicoll RA. Use-dependent increases in glutamate concentration activate presynaptic metabotropic glutamate receptors. Nature. 1997;385:630–634. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
