

Zbtb20 is essential for the specification of CA1 field identity in the developing hippocampus
- PMID: 20308569
- PMCID: PMC2851958
- DOI: 10.1073/pnas.0912315107
Zbtb20 is essential for the specification of CA1 field identity in the developing hippocampus
Abstract
The development of hippocampal circuitry depends on the proper assembly of correctly specified and fully differentiated hippocampal neurons. Little is known about factors that control the hippocampal specification. Here, we show that zinc finger protein Zbtb20 is essential for the specification of hippocampal CA1 field identity. We found that Zbtb20 expression was initially activated in the hippocampal anlage at the onset of corticogenesis, and persisted in immature hippocampal neurons. Targeted deletion of Zbtb20 in mice did not compromise the progenitor proliferation in the hippocampal and adjacent transitional ventricular zone, but led to the transformation of the hippocampal CA1 field into a transitional neocortex-like structure, as evidenced by cytoarchitectural, neuronal migration, and gene expression phenotypes. Correspondingly, the subiculum was ectopically located adjacent to the CA3 in mutant. Although the field identities of the mutant CA3 and dentate gyrus (DG) were largely maintained, their projections were severely impaired. The hippocampus of Zbtb20 null mice was reduced in size, and exhibited increased apoptotic cell death during postnatal development. Our data establish an essential role of Zbtb20 in the specification of CA1 field identity by repressing adjacent transitional neocortex-specific fate determination.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
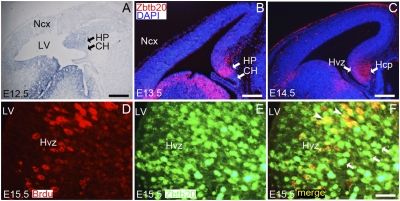
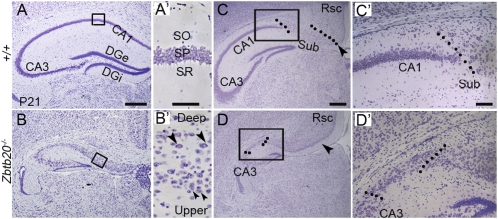
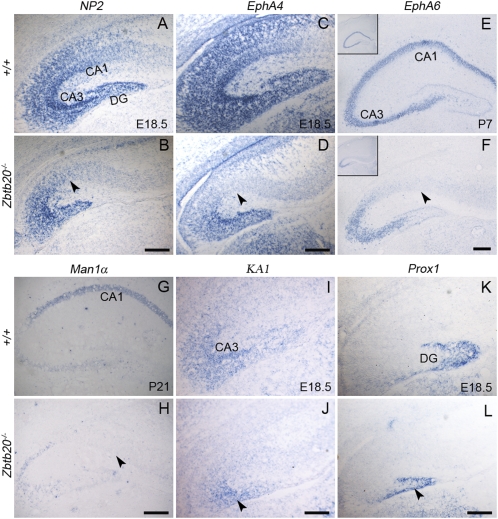
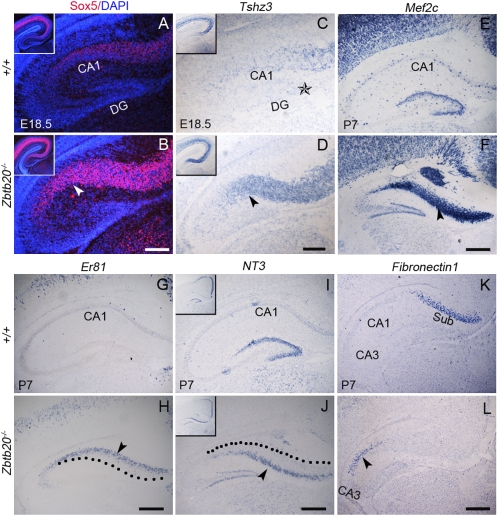
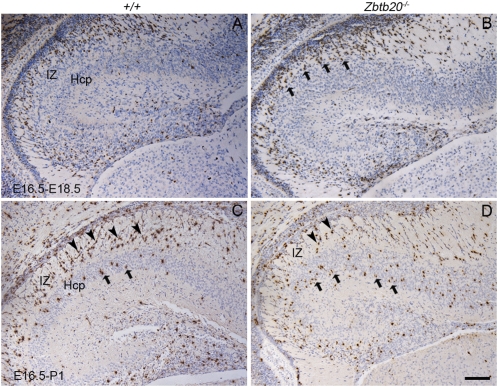
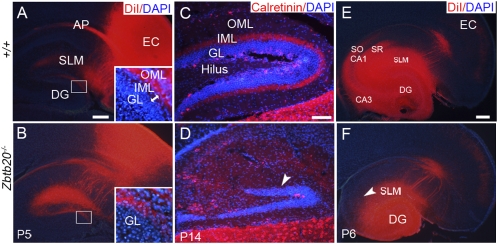
References
-
- Witter M, Amaral DG. In: The Rat Nervous System. Paxinos G, editor. San Diego: Elsevier Academic; 2004. pp. 635–704.
-
- Ishizuka N. Laminar organization of the pyramidal cell layer of the subiculum in the rat. J Comp Neurol. 2001;435:89–110. - PubMed
-
- Grove EA, Tole S, Limon J, Yip L, Ragsdale CW. The hem of the embryonic cerebral cortex is defined by the expression of multiple Wnt genes and is compromised in Gli3-deficient mice. Development. 1998;125:2315–2325. - PubMed
-
- Lee SM, Tole S, Grove E, McMahon AP. A local Wnt-3a signal is required for development of the mammalian hippocampus. Development. 2000;127:457–467. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
