

Systematic discovery of nonobvious human disease models through orthologous phenotypes
- PMID: 20308572
- PMCID: PMC2851946
- DOI: 10.1073/pnas.0910200107
Systematic discovery of nonobvious human disease models through orthologous phenotypes
Abstract
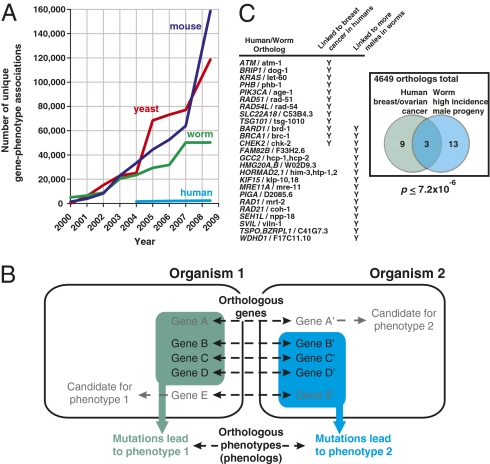
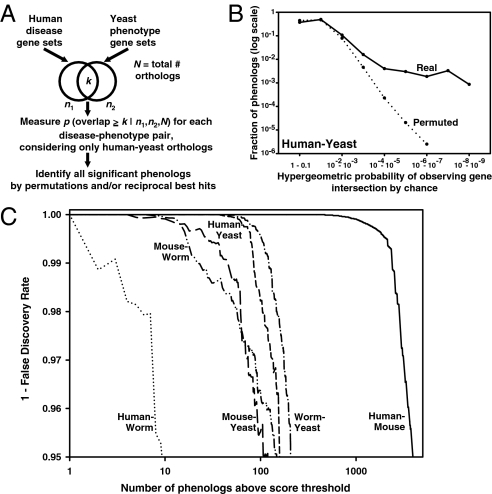
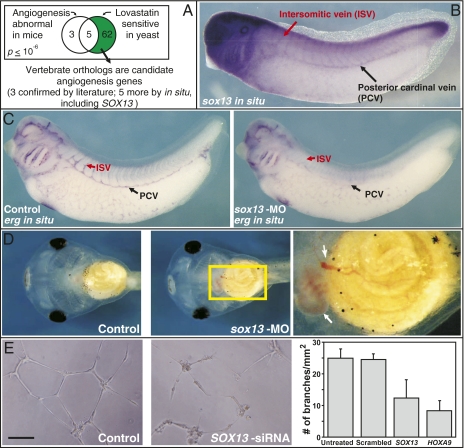
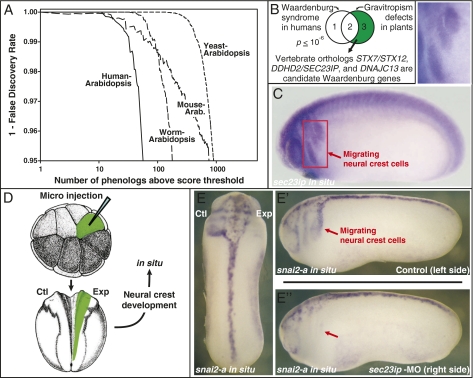
Biologists have long used model organisms to study human diseases, particularly when the model bears a close resemblance to the disease. We present a method that quantitatively and systematically identifies nonobvious equivalences between mutant phenotypes in different species, based on overlapping sets of orthologous genes from human, mouse, yeast, worm, and plant (212,542 gene-phenotype associations). These orthologous phenotypes, or phenologs, predict unique genes associated with diseases. Our method suggests a yeast model for angiogenesis defects, a worm model for breast cancer, mouse models of autism, and a plant model for the neural crest defects associated with Waardenburg syndrome, among others. Using these models, we show that SOX13 regulates angiogenesis, and that SEC23IP is a likely Waardenburg gene. Phenologs reveal functionally coherent, evolutionarily conserved gene networks-many predating the plant-animal divergence-capable of identifying candidate disease genes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dryja TP, et al. Homozygosity of chromosome 13 in retinoblastoma. N Engl J Med. 1984;310:550–553. - PubMed
-
- Lu X, Horvitz HR. lin-35 and lin-53, two genes that antagonize a C. elegans Ras pathway, encode proteins similar to Rb and its binding protein RbAp48. Cell. 1998;95:981–991. - PubMed
-
- Kassir Y, Rubin-Bejerano I, Mandel-Gutfreund Y. The Saccharomyces cerevisiae GSK-3 beta homologs. Curr Drug Targets. 2006;7:1455–1465. - PubMed
-
- Kim L, Kimmel AR. GSK3 at the edge: regulation of developmental specification and cell polarization. Curr Drug Targets. 2006;7:1411–1419. - PubMed
-
- Liu KJ, Arron JR, Stankunas K, Crabtree GR, Longaker MT. Chemical rescue of cleft palate and midline defects in conditional GSK-3beta mice. Nature. 2007;446:79–82. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
