

Do ferrets perceive relative pitch?
- PMID: 20329865
- PMCID: PMC2856516
- DOI: 10.1121/1.3290988
Do ferrets perceive relative pitch?
Abstract
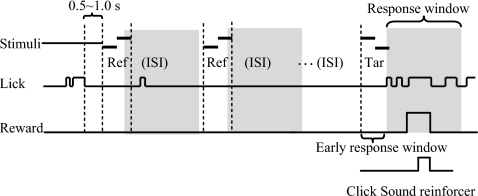
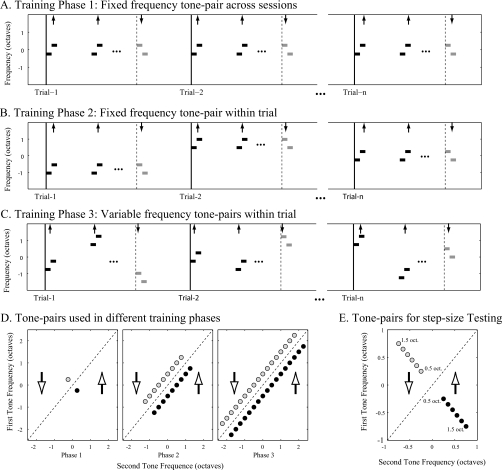
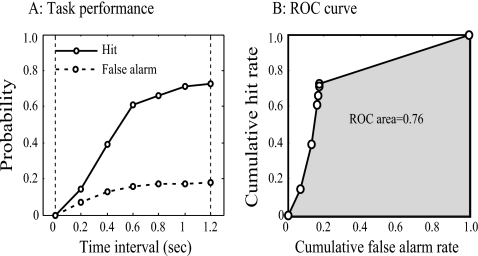
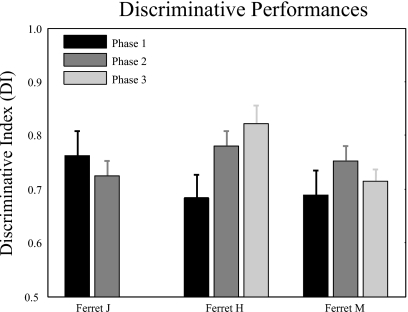
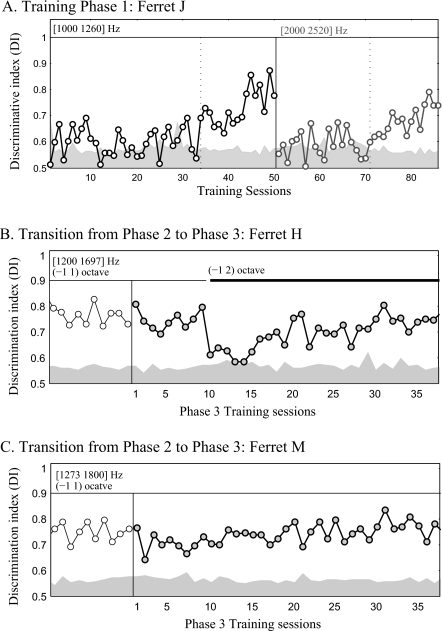
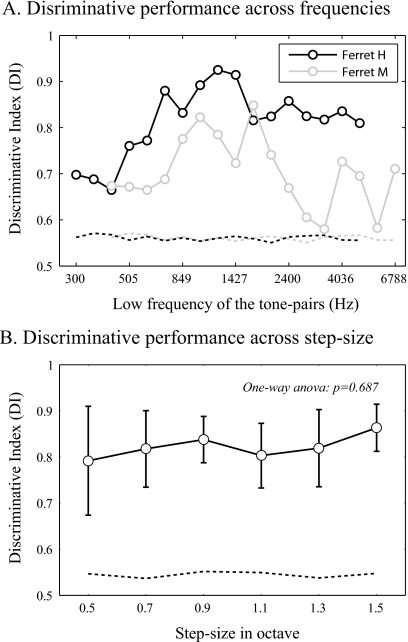
The existence of relative pitch perception in animals is difficult to demonstrate, since unlike humans, animals often attend to absolute rather than relative properties of sound elements. However, the results of the present study show that ferrets can be trained using relative pitch to discriminate two-tone sequences (rising vs. falling). Three ferrets were trained using a positive-reinforcement paradigm in which sequences of reference (one to five repeats) and target stimuli were presented, and animals were rewarded only when responding correctly to the target. The training procedure consisted of three training phases that successively shaped the ferrets to attend to relative pitch. In Phase-1 training, animals learned the basic task with sequences of invariant tone-pairs and could use absolute pitch information. During Phase-2 training, in order to emphasize relative cues, absolute pitch was varied each trial within a two-octave frequency range. In Phase-3 training, absolute pitch cues were removed, and only relative cue information was available to solve the task. Two ferrets successfully completed training on all three phases and achieved significant discriminative performance over the trained four-octave frequency range. These results suggest that ferrets can be trained to discern the relative pitch relationship of a sequence of tone-pairs independent of frequency.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources