

Ionizing radiation-induced long-term expression of senescence markers in mice is independent of p53 and immune status
- PMID: 20331441
- PMCID: PMC2894262
- DOI: 10.1111/j.1474-9726.2010.00567.x
Ionizing radiation-induced long-term expression of senescence markers in mice is independent of p53 and immune status
Abstract
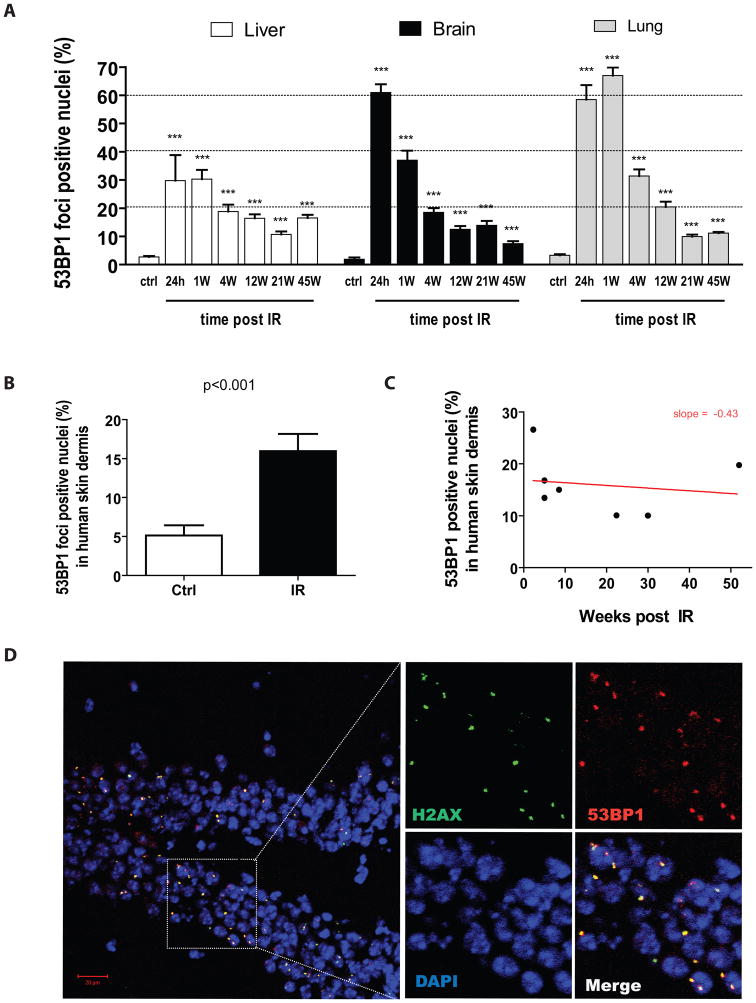
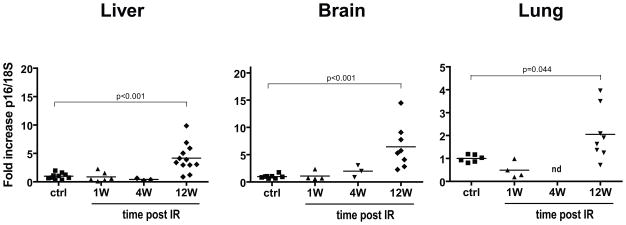
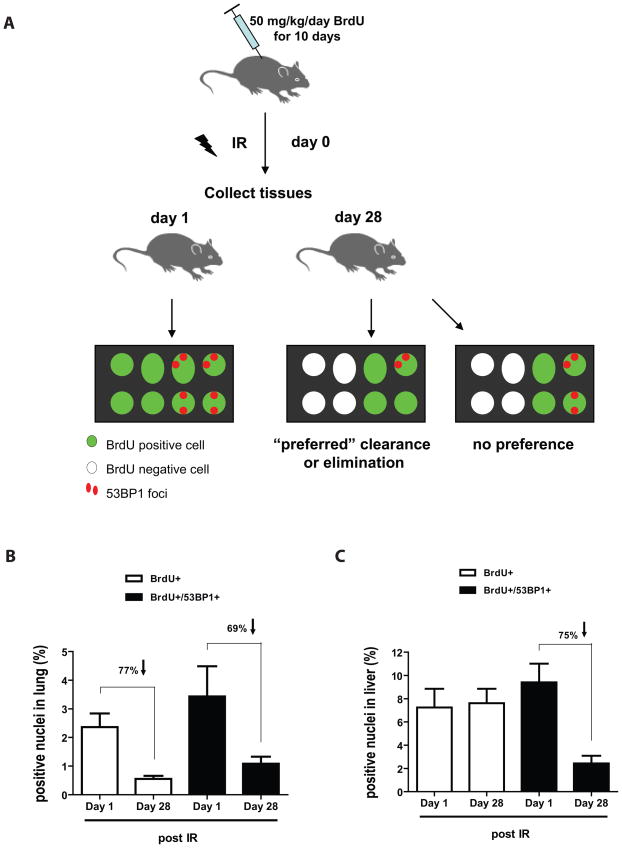
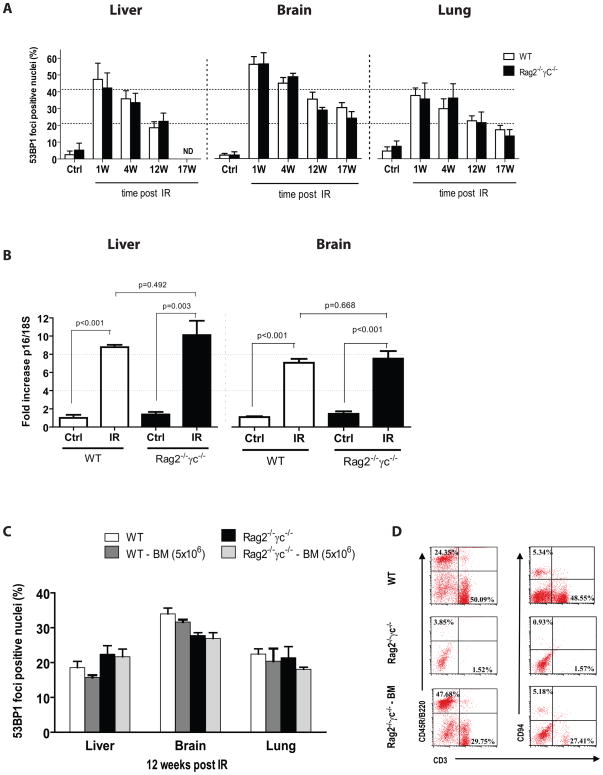
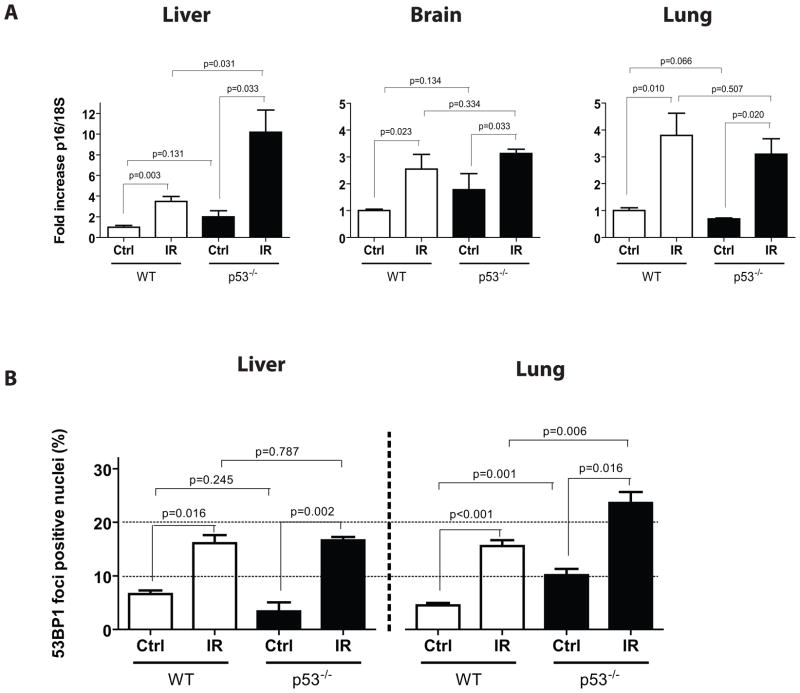
Exposure to IR has been shown to induce the formation of senescence markers, a phenotype that coincides with lifelong delayed repair and regeneration of irradiated tissues. We hypothesized that IR-induced senescence markers could persist long-term in vivo, possibly contributing to the permanent reduction in tissue functionality. Here, we show that mouse tissues exposed to a sublethal dose of IR display persistent (up to 45 weeks, the maximum time analyzed) DNA damage foci and increased p16(INK4a) expression, two hallmarks of cellular senescence and aging. BrdU-labeling experiments revealed that IR-induced damaged cells are preferentially eliminated, at least partially, in a tissue-dependent manner. Unexpectedly, the accumulation of damaged cells was found to occur independent from the DNA damage response modulator p53, and from an intact immune system, as their levels were similar in wild-type and Rag2(-/-) gammaC(-/-) mice, the latter being deficient in T, B, and NK cells. Together, our results provide compelling evidence that exposure to IR induces long-term expression of senescence markers in vivo, an effect that may contribute to the reduced tissue functionality observed in cancer survivors.
Conflict of interest statement
Figures
References
-
- Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, Vassiliou LV, Kolettas E, Niforou K, Zoumpourlis VC, Takaoka M, Nakagawa H, Tort F, Fugger K, Johansson F, Sehested M, Andersen CL, Dyrskjot L, Orntoft T, Lukas J, Kittas C, Helleday T, Halazonetis TD, Bartek J, Gorgoulis VG. Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature. 2006;444:633–637. - PubMed
-
- Begus-Nahrmann Y, Lechel A, Obenauf AC, Nalapareddy K, Peit E, Hoffmann E, Schlaudraff F, Liss B, Schirmacher P, Kestler H, Danenberg E, Barker N, Clevers H, Speicher MR, Rudolph KL. p53 deletion impairs clearance of chromosomal-instable stem cells in aging telomere-dysfunctional mice. Nat Genet. 2009;41:1138–1143. - PubMed
-
- Campisi J, d’Adda di Fagagna F. Cellular senescence: when bad things happen to good cells. Nat Rev Mol Cell Biol. 2007;8:729–740. - PubMed
-
- Chang C, Biedermann KA, Mezzina M, Brown JM. Characterization of the DNA double strand break repair defect in scid mice. Cancer Res. 1993;53:1244–1248. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
