

Bcl-XL and STAT3 mediate malignant actions of gamma-irradiation in lung cancer cells
- PMID: 20331635
- PMCID: PMC11159096
- DOI: 10.1111/j.1349-7006.2010.01552.x
Bcl-XL and STAT3 mediate malignant actions of gamma-irradiation in lung cancer cells
Abstract
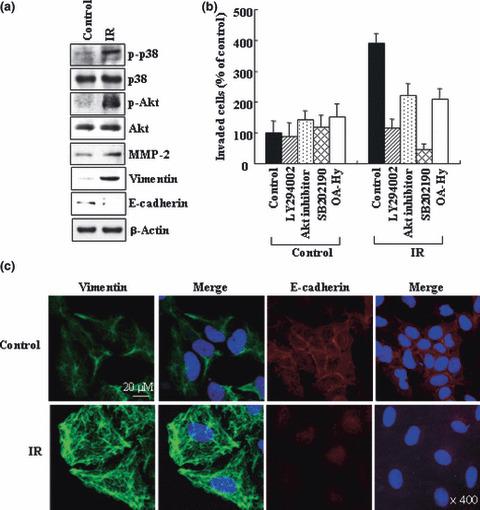
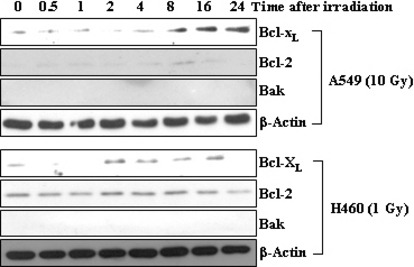
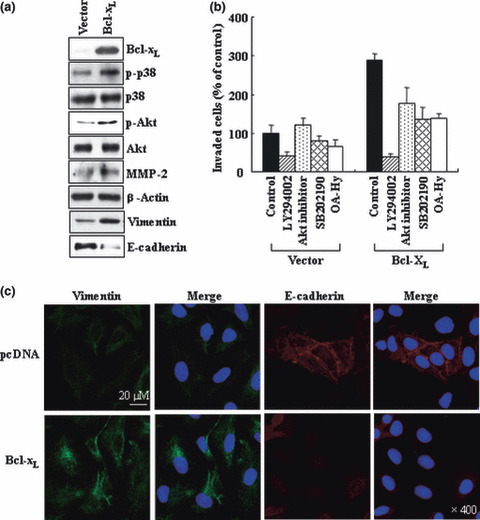
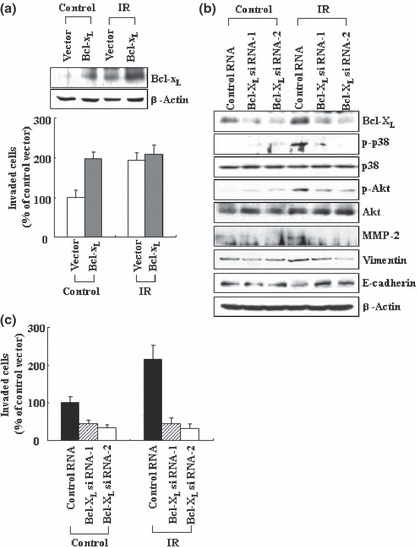
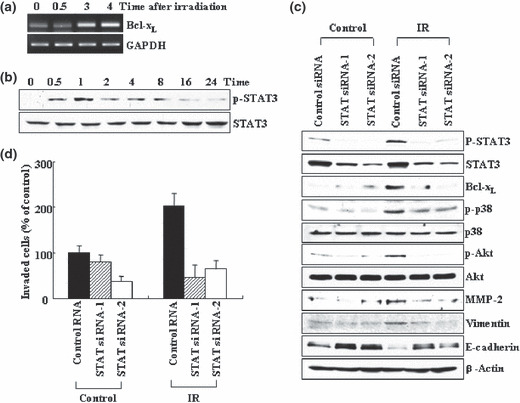
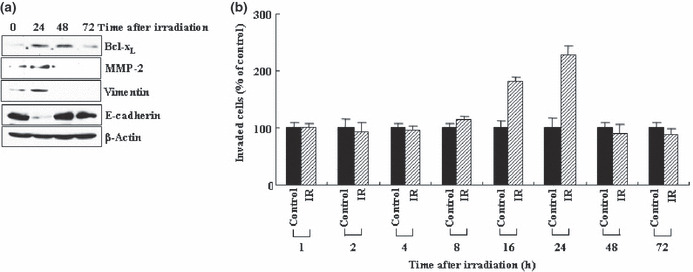
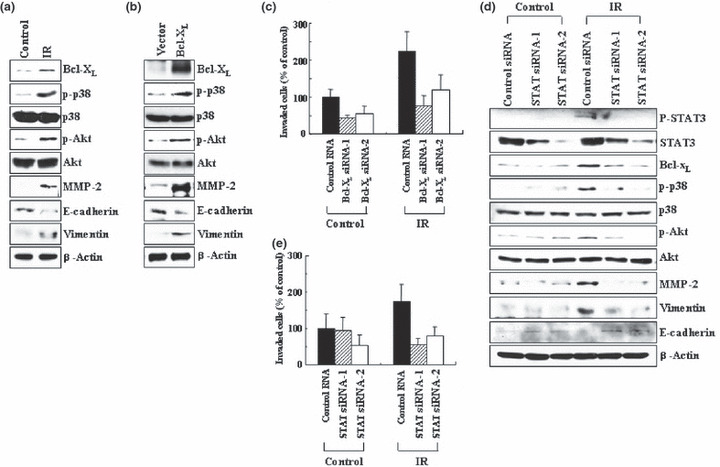
Previous reports suggest that, in addition to its therapeutic effects, ionizing radiation (IR) increases the invasiveness of surviving cancer cells. Here, we demonstrate that this activity of IR in lung cancer cells is mediated by a signaling pathway involving p38 kinase, phosphoinositide 3-kinase, Akt, and matrix metalloproteinase (MMP-2). The invasion-promoting doses of IR also increased and reduced the levels of vimentin and E-cadherin, respectively, both of which are markers for the epithelial-mesenchymal transition (EMT). Interestingly, all of these malignant actions of IR were mimicked by the overexpression of Bcl-X(L), a pro-survival member of the Bcl-2 family, in lung cancer cells. Moreover, both RNA and protein levels of Bcl-X(L) were elevated upon irradiation of the cells, and the prevention of this event using small-interfering RNAs of Bcl-X(L) reduced the ability of IR to promote invasion signals and EMT-associated events. This suggests that Bcl-X(L) functions as a signaling mediator of the malignant effects of IR. It was also demonstrated that IR enhances signal transducer and activator of transcription 3 (STAT3) phosphorylation, and the reduction of STAT3 levels via RNA interference prevented IR-induced Bcl-X(L) accumulation, and thus all the tested Bcl-X(L)-dependent events. Overall, the data suggest that IR induces Bcl-X(L) accumulation via STAT3, which then promotes cancer cell invasion and EMT-associated markers. Our findings demonstrate a novel function of Bcl-X(L) in cancer, and also advance our understanding of the malignant actions of IR significantly.
Figures
References
-
- Schmidt‐Ullrich RK. Molecular targets in radiation oncology. Oncogene 2003; 22: 5730–3. - PubMed
-
- Barcellos‐Hoff MH, Park C, Wright EG. Radiation and the microenvironment‐tumorigenesis and therapy. Nat Rev Cancer 2005; 5: 867–75. - PubMed
-
- Kumar P, Miller AI, Polverini PJ. Ap38 MAPK mediates γ‐irradiation‐induced endothelial cell apoptosis, and vascular endothelial growth factor protects endothelial cells through the phosphoinositide 3‐kinase‐Akt‐Bcl‐2 pathway. J Biol Chem 2004; 279: 43352–60. - PubMed
-
- Park CM, Park MJ, Kwak HJ et al. Ionizing radiation enhances matrix metalloproteinase‐2 secretion and invasion of glioma cells through Src/epidermal growth factor receptor‐mediated p38/Akt and phosphatidylinositol 3‐kinase/Akt signaling pathways. Cancer Res 2006; 66: 8511–9. - PubMed
-
- Cheng JH, Chou CH, Kuo ML, Hsieh CY. Radiation‐enhanced hepatocellular carinoma cell invasion with MMP‐9 expression through PI3K/Akt/NF‐κB signal transduction pathway. Oncogene 2006; 25: 7009–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
