

doi: 10.1242/jcs.056176.
Epub 2010 Mar 23.
Dishevelled-KSRP complex regulates Wnt signaling through post-transcriptional stabilization of beta-catenin mRNA
Affiliations
- PMID: 20332102
- PMCID: PMC2848118
- DOI: 10.1242/jcs.056176
Item in Clipboard
Dishevelled-KSRP complex regulates Wnt signaling through post-transcriptional stabilization of beta-catenin mRNA
J Cell Sci.
.
Abstract
Canonical Wnt/beta-catenin signaling is crucial during embryonic development. Upon Wnt stimulation, Dishevelled proteins relay the signal from upstream Frizzled receptors to downstream effectors. By using affinity purification followed by ion-trap mass spectrometry we identified K-homology splicing regulator protein (KSRP) as a novel Dishevelled-interacting protein. We show that KSRP negatively regulates Wnt/beta-catenin signaling at the level of post-transcriptional CTNNB1 (beta-catenin) mRNA stability. Thus, Dishevelled-KSRP complex operates in Wnt regulation of beta-catenin, functioning post-transcriptionally upon CTNNB1 mRNA stability.
Figures
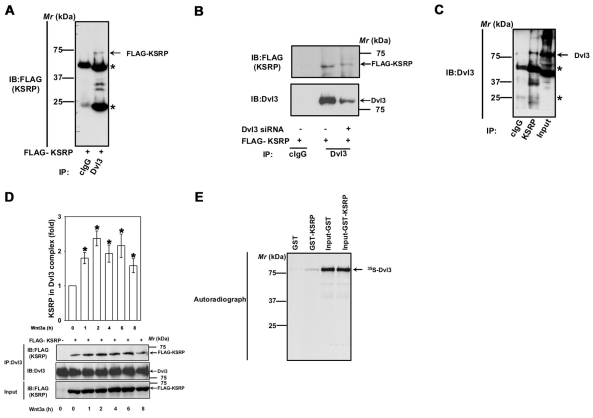
KSRP interacts with Dvl3. (A) F9 cells were transiently transfected with FLAG-KSRP for 24 hours followed by cell lysis and affinity pull-downs with either mouse control IgG or anti-Dvl3 mouse monoclonal antibody. Interaction of KSRP with Dvl3 was visualized by probing the blots with anti-FLAG antibody. Asterisks indicate the bands of immunoglobulin heavy and light chains. (B) F9 cells were treated with 100 nM of Dvl3 siRNA for 24 hours followed by transient expression of FLAG-KSRP for 24 hours followed by cell lysis and affinity pull-downs with either mouse control IgG or anti-Dvl3 mouse monoclonal antibody. Interaction of KSRP with Dvl3 was visualized by probing the blots with anti-FLAG antibody. (C) F9 cell lysates were immunoprecipitated with either rabbit control IgG or rabbit anti-KSRP polyclonal antibody and the interaction of KSRP with Dvl3 was visualized by probing the blots with anti-Dvl3 mouse monoclonal antibody. (D) F9 cells were transiently transfected with empty vector or FLAG-KSRP for 24 hours. The cells were then treated with Wnt3a (10 ng/ml) for indicated period of time followed by cell lysis and affinity pull-downs with anti-Dvl3 specific antibodies followed by immunoblotting with anti-FLAG antibodies. (E) To test the direct interaction of Dvl3 with KSRP, in vitro synthesized 35S-labeled Dvl3 was used in pull-down experiments with either GST- or GST-KSRP-Sepharose beads in the presence of 0.8% BSA. The interaction was visualized by SDS-PAGE and autoradiography. Representative blots of three independent experiments that proved highly reproducible are shown. *P<0.05 versus control (−Wnt3a).
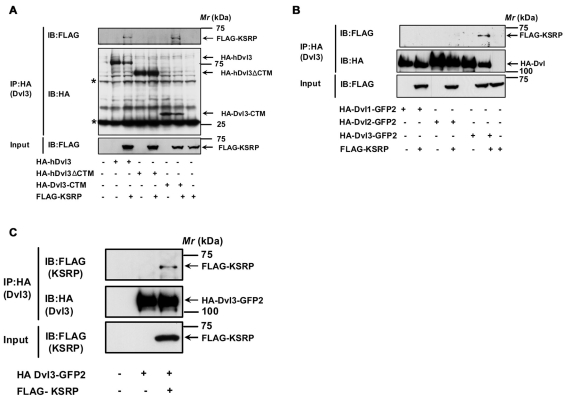
KSRP-Dvl3 interaction is mediated through the C-terminus of Dvl3. (A) F9 cells were treated with 100 nM Dvl3 siRNA for 24 hours followed by transient expression of either HA-hDvl3 or HA-hDvl3-ΔCTM (1-496) or HA-Dvl3-CTM (497-716) alone or together with FLAG-KSRP for 24 ho urs followed by cell lysis and affinity pull-downs with anti-HA-affinity matrix. Interaction of KSRP with Dvl3 or its mutants was visualized by probing the blots with anti-FLAG antibody. Asterisks indicate the bands of immunoglobulin hea vy and light chains. (B) F9 cells were transiently transfected with either HA-Dvl1-GFP2 or HA-Dvl2-GFP2 or HA-Dvl3-GFP2 alone or with FLAG-KSRP for 24 hours followed by cell lysis and affinity pull-downs with anti-HA-affinity matrix. Interaction of KSRP with Dvl isoforms was visualized by probing the blots with anti-FLAG antibody. (C) HEK293 cells were transiently transfected with HA-Dvl3-GFP2 alone or with FLAG-KSRP for 24 hours followed by cell lysis and affinity pull-downs with anti-HA-affinity matrix. Interaction of KSRP with Dvl3 was visualized by probing the blots with anti-FLAG antibody. Representative blots of three independent experiments that proved highly reproducible are displayed.
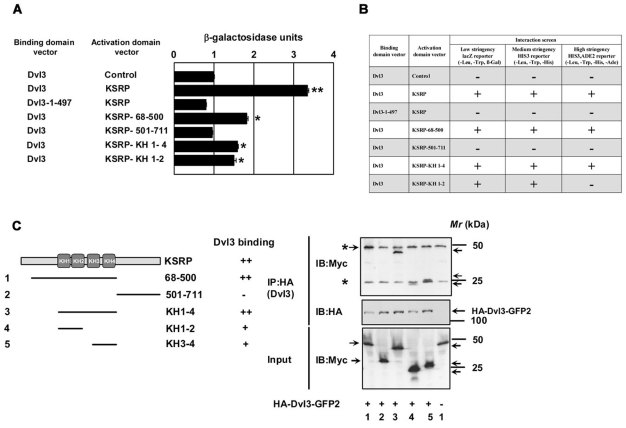
Dvl3 interacts with KSRP via N-terminal KH domains. Yeast-two hybrid assays were performed using full-length Dvl3 and deletion mutant of Dvl3 (1-497) in the DNA-binding domain vector and the full-length KSRP and the deletion mutants of KSRP in the activation domain vector. (A) To test for positive protein-protein interaction, quantitative β-galactosidase assays were performed. The results from liquid ONPG assays (displayed in β-galactosidase units) are represented in the graph. Dvl3-pGAD-AD (control) was used as a control for self-activation. The data represent mean ± s.e.m. obtained from two independent experiments performed in triplicate. *P<0.05; **P<0.01 versus control (Dvl3-pGAD-AD control). (B) Growth of yeast strains expressing fusion proteins on selective medium were also tested for studying protein-protein interactions. All the yeast cultures used for liquid β-galactosidase assays were normalized to similar OD600 densities and spotted onto selection plates [synthetic minimal medium lacking leucine, tryptophan and histidine (medium stringency)] or [synthetic minimal medium lacking leucine, tryptophan, histidine and adenine (high stringency)]. Yeast transformants that survive on the selection plates (medium and high stringency) were considered positive for an interaction, as shown in B. (C) To test the interaction of Dvl3 and the deletion mutants of KSRP in vivo, F9 cells were transiently transfected with the indicated Myc-tagged KSRP deletion constructs (left panel) along with HA-Dvl3-GFP2 for 24 hours followed by cell lysis and affinity pull-downs with anti-HA affinity matrix. Interaction of Dvl3 with deletion mutants of KSRP was visualized by probing the blots with anti-Myc antibody. The immunoreactive band for the N-terminus of KSRP (68-500) protein was observed to migrate along with the immunoglobulin heavy chain protein (as revealed by the darker intensity in lane 1 when compared with others). Asterisks indicate the bands of immunoglobulin heavy and light chains. Representative blots of two independent experiments that proved highly reproducible are displayed.

KSRP negatively regulates Wnt/β-catenin signaling. (A) F9 cells were treated with either control siRNAs (100 nM) or siRNAs specific to Khsrp, Dvl2 or Dvl3 (100 nM) for 48 hours and the Ctnnb1 and Ppia (cyclophilin A) mRNA levels were quantified using quantitative PCR. The data represent Ctnnb1 mRNA normalized to the Ppia (cyclophilin A) mRNA levels (mean values ± s.e.m.) from three independent experiments whose results were in high agreement. F9 cells were treated with either control siRNAs (100 nM) or siRNAs specific for mouse Khsrp (100 nM) for 48 hours and the lysates were assayed either for cytosolic β-catenin levels (B) or Lef/Tcf-sensitive transcription (C). Upper panel displays mean values ± s.e.m. from three independent experiments; the lower panel displays representative blots. (D) F9 cells were treated with control siRNAs (100 nM) or siRNAs specific for mouse Khsrp (100 nM) for 48 hours before stimulation with Wnt3a for 4 days. Subsequently, formation of primitive endoderm was assayed by immunostaining the cells with anti-cytokeratin endo A antibody (TROMA-1), a hallmark of PE formation. Typical phase-contrast images (PC) and the indirect immunofluorescence images (IIF) are shown from a single experiment, representative of two independent experiments. F9 cells (E) or HEK293 cells (F) were transfected with FLAG-KSRP for 24 hours and the lysates were assayed for cytosolic β-catenin stabilization. (G) F9 cells were transfected with indicated amounts of FLAG-KSRP for 24 hours and the lysates were assayed for Lef/Tcf-sensitive transcription after stimulation with Wnt3a for 7 hours. (H) F9 cells were transfected with HA-Dvl3-GFP2 alone or together with increasing amounts of FLAG-KSRP for 24 hours and the lysates were assayed for Lef/Tcf-sensitive transcription. (I) F9 cells were transfected with indicated amounts of an unrelated protein, mouse S-Phase kinase associated protein 1A (mSKP1A) for 24 hours and the lysates were assayed for Lef/Tcf-sensitive transcription. The data represents mean values ± s.e.m. from three independent experiments *P<0.05; **P<0.01 versus control (−Wnt3a). #P<0.05; ##P<0.01 versus control (+Wnt3a).
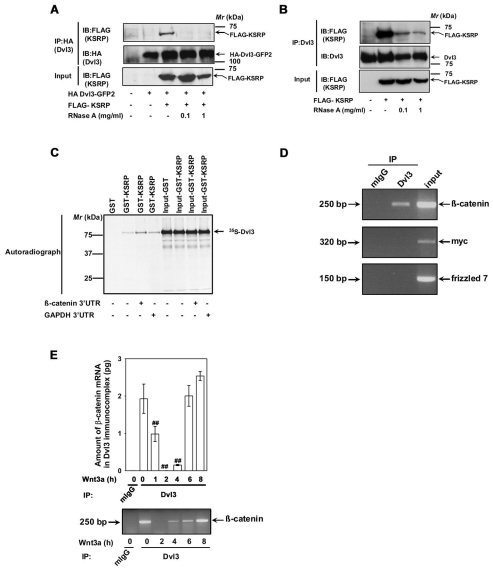
Dvl3-KSRP interaction is RNA dependent and Dvl3 complex harbors Ctnnb1 mRNA. (A) F9 cells were co-transfected with HA-Dvl3-GFP2 and FLAG-KSRP for 24 hours followed by cell lysis. The lysates were then incubated without or with indicated amounts of RNaseA at room temperature for 10 minutes followed by affinity pull-downs with anti-HA affinity matrix. Interaction of KSRP with exogenous Dvl3 was probed by immunoblotting with anti-FLAG antibodies. (B) F9 cells were transfected with FLAG-KSRP for 24 hours followed by cell lysis. The lysates were then incubated without or with indicated amounts of RNaseA at room temperature for 10 minutes followed by affinity pull-downs with anti-Dvl3 antibodies. Interaction of KSRP with endogenous Dvl3 was probed by immunoblotting with anti-FLAG antibodies. (C) To test whether the in vitro interaction of KSRP and Dvl3 is also RNA dependent, in vitro synthesized 35S-labeled Dvl3 was used in pull-down experiments with either GST or GST-KSRP-Sepharose beads in the absence or presence (5 μg) of either 3′-UTR of Ctnnb1 or Gapdh. The interaction was visualized by SDS-PAGE and autoradiography. (D) RNA immunoprecipitation assay was performed on F9 cell lysates with either control mouse IgG or anti-Dvl3 antibodies. The RNA isolated from the immunoprecipitates was analyzed by RT-PCR with primers specific for Ctnnb1, Myc or Fzd7. Representative gel of two independent experiments that proved highly reproducible is displayed. (E) F9 cells were treated with Wnt3a (10 ng/ml) for indicated periods of time and RNA immunoprecipitation assay was performed with either control mouse IgG or anti-Dvl3 antibodies. The RNA isolated from the immunoprecipitates was analyzed by quantitative real-time PCR with β-catenin specific primers. Representative blots of two independent experiments that proved highly reproducible were displayed. The data represent mean values ± s.e.m. from two independent experiments whose results were in high agreement. In the lower panel, a representative gel is displayed. ##P<0.01 versus the control (−Wnt3a).
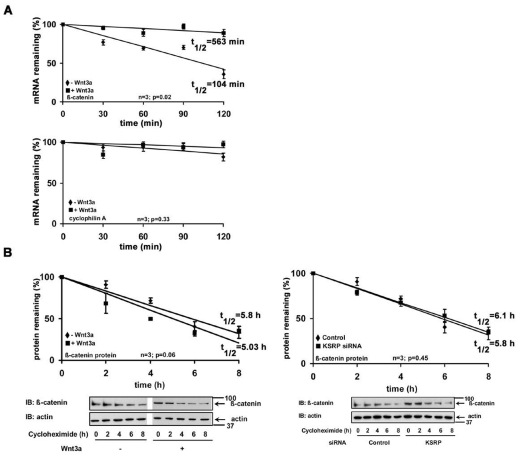
Ctnnb1 mRNA has a short half-life. (A) Serum-starved F9 cells were treated without or with Wnt3a (10 ng/ml) for 1 hour, followed by treatment with actinomycin D (5 μg/ml) for indicated periods of time. Ctnnb1 and Ppia (cyclophilin A) mRNA levels were quantified using quantitative PCR. The data represents mean values ± s.e.m. from three independent experiments whose results were in high agreement. (B) β-catenin half-life was estimated either in F9 cells or F9 cells previously treated with Khsrp siRNAs, by treating the cells with cycloheximide (100 μg/ml) for indicated periods of time, followed by immunoblotting and densitometric scanning. β-catenin levels were normalized by the actin levels. The data represents mean values ± s.e.m. from three independent experiments whose results were in high agreement.

Wnt3a-induced β-catenin accumulation is the result of post-transcriptional stabilization of Ctnnb1 mRNA. (A) Confluent F9 cells were treated with vehicle (DMSO), actinomycin D (Act D, 5 μg/ml) or cycloheximide (CHX, 100 μg/ml) for 1 hour followed by stimulation with Wnt3a for 7 hours. Lef/Tcf-sensitive transcription was determined. The data represent mean values ± s.e.m. from three separate experiments performed in triplicates. (B) Confluent F9 cells were treated with vehicle (DMSO), actinomycin D (5 μg/ml) or cycloheximide (100 μg/ml) for 1 hour before addition of Wnt3a for indicated periods of time. After stimulation, the lysates were collected and cytosolic β-catenin levels were assayed. Upper panel displays mean values ± s.e.m. obtained from three independent experiments; the lower panel displays representative blots. **P<0.01 versus control (−Wnt3a); #P<0.05; ##P<0.01 versus the control (+Wnt3a).
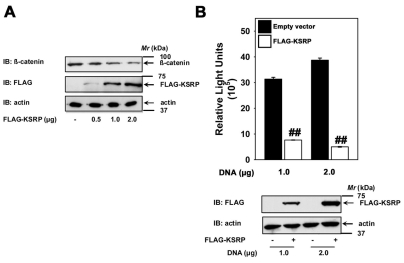
KSRP overexpression effects β-catenin signaling in colon carcinoma cells. SW480 cells (2.5×105 cells/well in a 12-well cell culture dish) were transfected with indicated amounts of either empty vector or FLAG-KSRP plasmid for 24 hours using a Fugene transfection reagent (Roche applied life sciences) to DNA ratio of 5:2. The lysates were then assayed either for cytosolic β-catenin levels (A) or Lef/Tcf-sensitive transcription (B). Upper panel in B displays mean values ± s.e.m. obtained from three independent experiments; the lower panel displays representative blots. For the β-catenin assay representative blots of two independent experiments that proved highly reproducible are displayed. ##P<0.01 versus the control (empty vector).
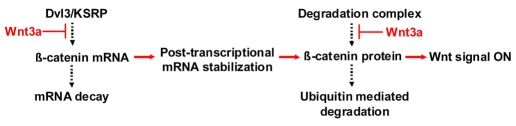
Regulation of β-catenin levels in canonical Wnt/β-catenin signaling pathway. Basal levels of β-catenin are regulated at the mRNA level by Dvl3-KSRP complex that promotes de-stabilization of CTNNB1 mRNA, and at the protein level by GSK3β-mediated phosphorylation that promotes degradation of β-catenin. Wnt stimulation contributes to rapid accumulation of β-catenin by opposing the ability of the Dvl3-KSRP complex to promote destabilization of CTNNB1 mRNA and the degradation of β-catenin mediated by GSK3β phosphorylation.
Similar articles
-
Dishevelled-2 docks and activates Src in a Wnt-dependent manner.J Cell Sci. 2009 Dec 15;122(Pt 24):4439-51. doi: 10.1242/jcs.051847. Epub 2009 Nov 17. J Cell Sci. 2009. PMID: 19920076 Free PMC article.
-
A novel Wilms tumor 1 (WT1) target gene negatively regulates the WNT signaling pathway.J Biol Chem. 2010 May 7;285(19):14585-93. doi: 10.1074/jbc.M109.094334. Epub 2010 Mar 10. J Biol Chem. 2010. PMID: 20220130 Free PMC article.
-
Loss of the tumor suppressor CYLD enhances Wnt/beta-catenin signaling through K63-linked ubiquitination of Dvl.Mol Cell. 2010 Mar 12;37(5):607-19. doi: 10.1016/j.molcel.2010.01.035. Mol Cell. 2010. PMID: 20227366
-
Dishevelled: The hub of Wnt signaling.Cell Signal. 2010 May;22(5):717-27. doi: 10.1016/j.cellsig.2009.11.021. Epub 2009 Dec 13. Cell Signal. 2010. PMID: 20006983 Review.
-
New steps in the Wnt/beta-catenin signal transduction pathway.Recent Prog Horm Res. 2000;55:225-36. Recent Prog Horm Res. 2000. PMID: 11036939 Review.
Cited by
-
KHSRP-bound small nucleolar RNAs associate with promotion of cell invasiveness and metastasis of pancreatic cancer.Oncotarget. 2020 Jan 14;11(2):131-147. doi: 10.18632/oncotarget.27413. eCollection 2020 Jan 14. Oncotarget. 2020. PMID: 32010427 Free PMC article.
-
miR-3619-3p promotes papillary thyroid carcinoma progression via Wnt/β-catenin pathway.Ann Transl Med. 2019 Nov;7(22):643. doi: 10.21037/atm.2019.10.71. Ann Transl Med. 2019. PMID: 31930044 Free PMC article.
-
KSRP-mediated Wnt/β-catenin activation promotes follicular thyroid cancer progression and stemness.Br J Cancer. 2025 Aug 23. doi: 10.1038/s41416-025-03142-x. Online ahead of print. Br J Cancer. 2025. PMID: 40849355
-
Proteasome machinery is instrumental in a common gain-of-function program of the p53 missense mutants in cancer.Nat Cell Biol. 2016 Aug;18(8):897-909. doi: 10.1038/ncb3380. Epub 2016 Jun 27. Nat Cell Biol. 2016. PMID: 27347849
-
The RNA-Binding Protein KSRP Modulates Cytokine Expression of CD4+ T Cells.J Immunol Res. 2019 Aug 14;2019:4726532. doi: 10.1155/2019/4726532. eCollection 2019. J Immunol Res. 2019. PMID: 31511826 Free PMC article.
References
-
- Aghib D. F., McCrea P. D. (1995). The E-cadherin complex contains the src substrate p120. Exp. Cell Res. 218, 359-369 - PubMed
-
- Behrens J., von Kries J. P., Kuhl M., Bruhn L., Wedlich D., Grosschedl R., Birchmeier W. (1996). Functional interaction of beta-catenin with the transcription factor LEF-1. Nature 382, 638-642 - PubMed
-
- Bikkavilli R. K., Feigin M. E., Malbon C. C. (2008). G alpha o mediates WNT-JNK signaling through dishevelled 1 and 3, RhoA family members, and MEKK 1 and 4 in mammalian cells. J. Cell Sci. 121, 234-245 - PubMed
-
- Boutros M., Paricio N., Strutt D. I., Mlodzik M. (1998). Dishevelled activates JNK and discriminates between JNK pathways in planar polarity and wingless signaling. Cell 94, 109-118 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
