

Information processing without brains--the power of intercellular regulators in plants
- PMID: 20332147
- PMCID: PMC2847462
- DOI: 10.1242/dev.034868
Information processing without brains--the power of intercellular regulators in plants
Abstract
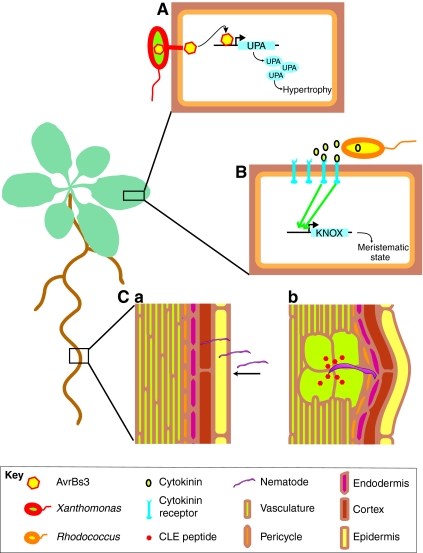
Plants exhibit different developmental strategies than animals; these are characterized by a tight linkage between environmental conditions and development. As plants have neither specialized sensory organs nor a nervous system, intercellular regulators are essential for their development. Recently, major advances have been made in understanding how intercellular regulation is achieved in plants on a molecular level. Plants use a variety of molecules for intercellular regulation: hormones are used as systemic signals that are interpreted at the individual-cell level; receptor peptide-ligand systems regulate local homeostasis; moving transcriptional regulators act in a switch-like manner over small and large distances. Together, these mechanisms coherently coordinate developmental decisions with resource allocation and growth.
Figures
Similar articles
-
Plant hormones and nutrient signaling.Plant Mol Biol. 2009 Mar;69(4):361-73. doi: 10.1007/s11103-008-9380-y. Epub 2008 Aug 9. Plant Mol Biol. 2009. PMID: 18688730 Review.
-
Auxin as a model for the integration of hormonal signal processing and transduction.Mol Plant. 2008 Mar;1(2):229-37. doi: 10.1093/mp/ssn006. Mol Plant. 2008. PMID: 19825535 Review.
-
Hormone interactions during vascular development.Plant Mol Biol. 2009 Mar;69(4):347-60. doi: 10.1007/s11103-008-9374-9. Epub 2008 Jul 25. Plant Mol Biol. 2009. PMID: 18654740 Review.
-
Molecular genetic studies confirm the role of brassinosteroids in plant growth and development.Plant J. 1996 Jul;10(1):1-8. doi: 10.1046/j.1365-313x.1996.10010001.x. Plant J. 1996. PMID: 8758975 Review. No abstract available.
-
Genomic analysis of DELLA protein activity.Plant Cell Physiol. 2013 Aug;54(8):1229-37. doi: 10.1093/pcp/pct082. Epub 2013 Jun 18. Plant Cell Physiol. 2013. PMID: 23784221 Review.
Cited by
-
PlantPhoneDB: A manually curated pan-plant database of ligand-receptor pairs infers cell-cell communication.Plant Biotechnol J. 2022 Nov;20(11):2123-2134. doi: 10.1111/pbi.13893. Epub 2022 Jul 30. Plant Biotechnol J. 2022. PMID: 35842742 Free PMC article.
-
Gut Microbiota and Neuroplasticity.Cells. 2021 Aug 13;10(8):2084. doi: 10.3390/cells10082084. Cells. 2021. PMID: 34440854 Free PMC article. Review.
-
Phytoplasma effector SAP54 induces indeterminate leaf-like flower development in Arabidopsis plants.Plant Physiol. 2011 Oct;157(2):831-41. doi: 10.1104/pp.111.181586. Epub 2011 Aug 17. Plant Physiol. 2011. PMID: 21849514 Free PMC article.
-
Identification and Expression Analysis of Phosphatidylinositol Transfer Proteins Genes in Rice.Plants (Basel). 2023 May 26;12(11):2122. doi: 10.3390/plants12112122. Plants (Basel). 2023. PMID: 37299101 Free PMC article.
-
TERMINAL FLOWER1 Functions as a Mobile Transcriptional Cofactor in the Shoot Apical Meristem.Plant Physiol. 2020 Apr;182(4):2081-2095. doi: 10.1104/pp.19.00867. Epub 2020 Jan 29. Plant Physiol. 2020. PMID: 31996406 Free PMC article.
References
-
- Abe M., Kobayashi Y., Yamamoto S., Daimon Y., Yamaguchi A., Ikeda Y., Ichinoki H., Notaguchi M., Goto K., Araki T. (2005). FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309, 1052-1056 - PubMed
-
- Akiyama K., Matsuzaki K., Hayashi H. (2005). Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435, 824-827 - PubMed
-
- An H., Roussot C., Suarez-Lopez P., Corbesier L., Vincent C., Pineiro M., Hepworth S., Mouradov A., Justin S., Turnbull C., et al. (2004). CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development 131, 3615-3626 - PubMed
-
- Angra R., Mandahar C. L. (1991). Pathogenesis of barley leaves by Helminthosporium teres I: Green island formation and the possible involvement of cytokinins. Mycopathologia 114, 21-27
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
