

On the origin and evolution of vertebrate olfactory receptor genes: comparative genome analysis among 23 chordate species
- PMID: 20333175
- PMCID: PMC2817399
- DOI: 10.1093/gbe/evp003
On the origin and evolution of vertebrate olfactory receptor genes: comparative genome analysis among 23 chordate species
Abstract
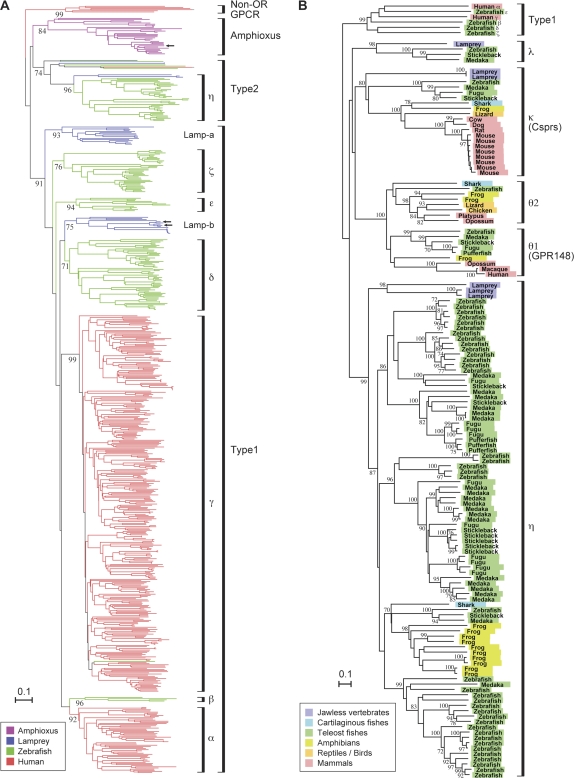
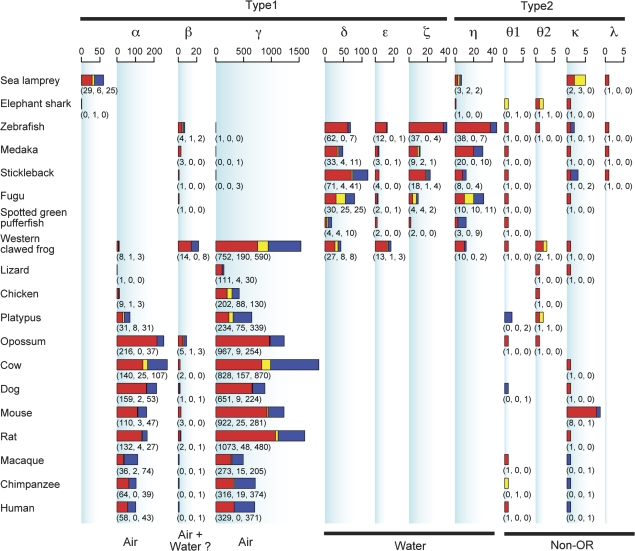
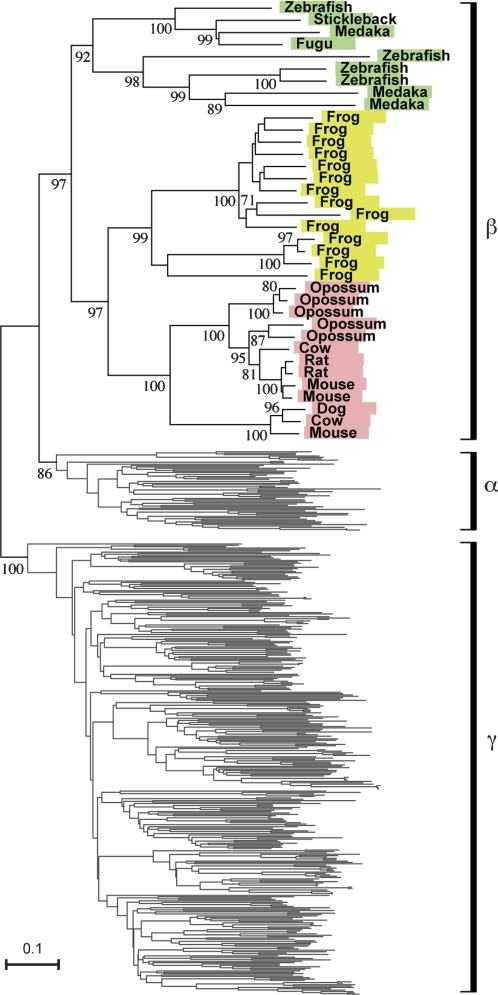
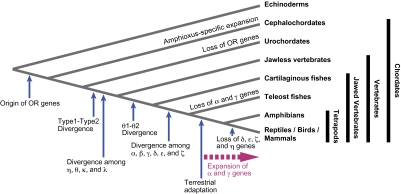
Olfaction is a primitive sense in organisms. Both vertebrates and insects have receptors for detecting odor molecules in the environment, but the evolutionary origins of these genes are different. Among studied vertebrates, mammals have approximately 1,000 olfactory receptor (OR) genes, whereas teleost fishes have much smaller (approximately 100) numbers of OR genes. To investigate the origin and evolution of vertebrate OR genes, I attempted to determine near-complete OR gene repertoires by searching whole-genome sequences of 14 nonmammalian chordates, including cephalochordates (amphioxus), urochordates (ascidian and larvacean), and vertebrates (sea lamprey, elephant shark, five teleost fishes, frog, lizard, and chicken), followed by a large-scale phylogenetic analysis in conjunction with mammalian OR genes identified from nine species. This analysis showed that the amphioxus has >30 vertebrate-type OR genes though it lacks distinctive olfactory organs, whereas all OR genes appear to have been lost in the urochordate lineage. Some groups of genes (theta, kappa, and lambda) that are phylogenetically nested within vertebrate OR genes showed few gene gains and losses, which is in sharp contrast to the evolutionary pattern of OR genes, suggesting that they are actually non-OR genes. Moreover, the analysis demonstrated a great difference in OR gene repertoires between aquatic and terrestrial vertebrates, reflecting the necessity for the detection of water-soluble and airborne odorants, respectively. However, a minor group (beta) of genes that are atypically present in both aquatic and terrestrial vertebrates was also found. These findings should provide a critical foundation for further physiological, behavioral, and evolutionary studies of olfaction in various organisms.
Keywords: amphioxus; chordate; molecular evolution; multigene family; olfactory receptor; vertebrate.
Figures
Similar articles
-
Evolution of developmental roles of Pax2/5/8 paralogs after independent duplication in urochordate and vertebrate lineages.BMC Biol. 2008 Aug 22;6:35. doi: 10.1186/1741-7007-6-35. BMC Biol. 2008. PMID: 18721460 Free PMC article.
-
Evolutionary dynamics of olfactory receptor genes in chordates: interaction between environments and genomic contents.Hum Genomics. 2009 Dec;4(2):107-18. doi: 10.1186/1479-7364-4-2-107. Hum Genomics. 2009. PMID: 20038498 Free PMC article. Review.
-
Olfactory receptor multigene family in vertebrates: from the viewpoint of evolutionary genomics.Curr Genomics. 2012 Apr;13(2):103-14. doi: 10.2174/138920212799860706. Curr Genomics. 2012. PMID: 23024602 Free PMC article.
-
Characterization of the neurohypophysial hormone gene loci in elephant shark and the Japanese lamprey: origin of the vertebrate neurohypophysial hormone genes.BMC Evol Biol. 2009 Feb 26;9:47. doi: 10.1186/1471-2148-9-47. BMC Evol Biol. 2009. PMID: 19243634 Free PMC article.
-
Development, metamorphosis, morphology, and diversity: The evolution of chordate muscles and the origin of vertebrates.Dev Dyn. 2015 Sep;244(9):1046-1057. doi: 10.1002/dvdy.24245. Epub 2015 Jul 14. Dev Dyn. 2015. Retraction in: Dev Dyn. 2015 Sep;244(9):1179. doi: 10.1002/dvdy.24236. PMID: 26095777 Retracted. Review.
Cited by
-
The Remarkable Diversity of Vertebrate Bitter Taste Receptors: Recent Advances in Genomic and Functional Studies.Int J Mol Sci. 2024 Nov 25;25(23):12654. doi: 10.3390/ijms252312654. Int J Mol Sci. 2024. PMID: 39684366 Free PMC article. Review.
-
Expression of Olfactory Sense-Related Genes in Chicken Olfactory Epithelium.J Poult Sci. 2025 Jul 18;62:2025019. doi: 10.2141/jpsa.2025019. eCollection 2025. J Poult Sci. 2025. PMID: 40689228 Free PMC article.
-
Chemosensory Receptors in Vertebrates: Structure and Computational Modeling Insights.Int J Mol Sci. 2025 Jul 10;26(14):6605. doi: 10.3390/ijms26146605. Int J Mol Sci. 2025. PMID: 40724854 Free PMC article. Review.
-
Whole genome sequencing of a snailfish from the Yap Trench (~7,000 m) clarifies the molecular mechanisms underlying adaptation to the deep sea.PLoS Genet. 2021 May 13;17(5):e1009530. doi: 10.1371/journal.pgen.1009530. eCollection 2021 May. PLoS Genet. 2021. PMID: 33983934 Free PMC article.
-
Massive Loss of Olfactory Receptors But Not Trace Amine-Associated Receptors in the World's Deepest-Living Fish (Pseudoliparis swirei).Genes (Basel). 2019 Nov 8;10(11):910. doi: 10.3390/genes10110910. Genes (Basel). 2019. PMID: 31717379 Free PMC article.
References
-
- Aparicio S, et al. Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science. 2002;297:1301–1310. - PubMed
-
- Berghard A, Dryer L. A novel family of ancient vertebrate odorant receptors. J Neurobiol. 1998;37:383–392. - PubMed
-
- Brusca RC, Brusca GJ. Invertebrates. 2nd ed. Sunderland (MA): Sinauer Associates; 2003.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
