

Phylogenomics of sterol synthesis: insights into the origin, evolution, and diversity of a key eukaryotic feature
- PMID: 20333205
- PMCID: PMC2817430
- DOI: 10.1093/gbe/evp036
Phylogenomics of sterol synthesis: insights into the origin, evolution, and diversity of a key eukaryotic feature
Abstract
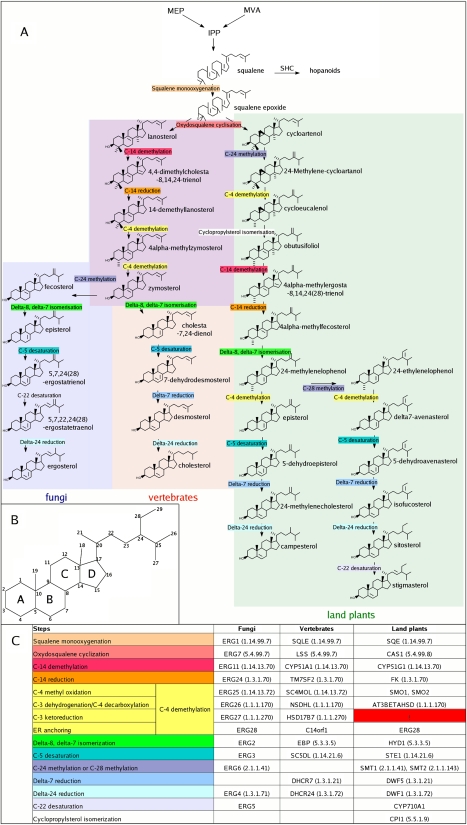
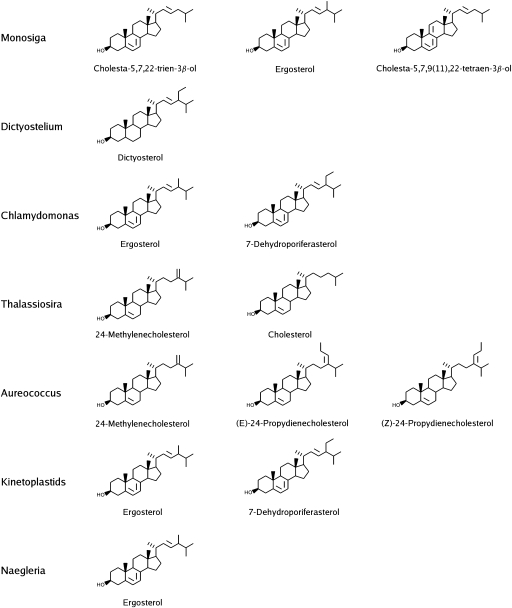
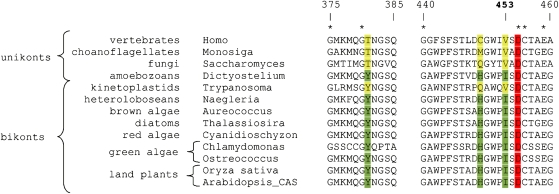
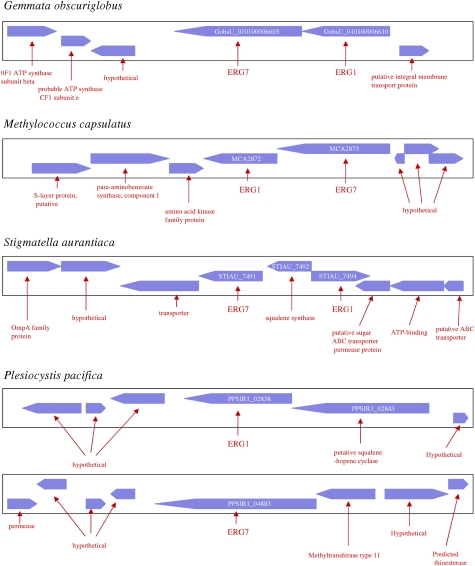
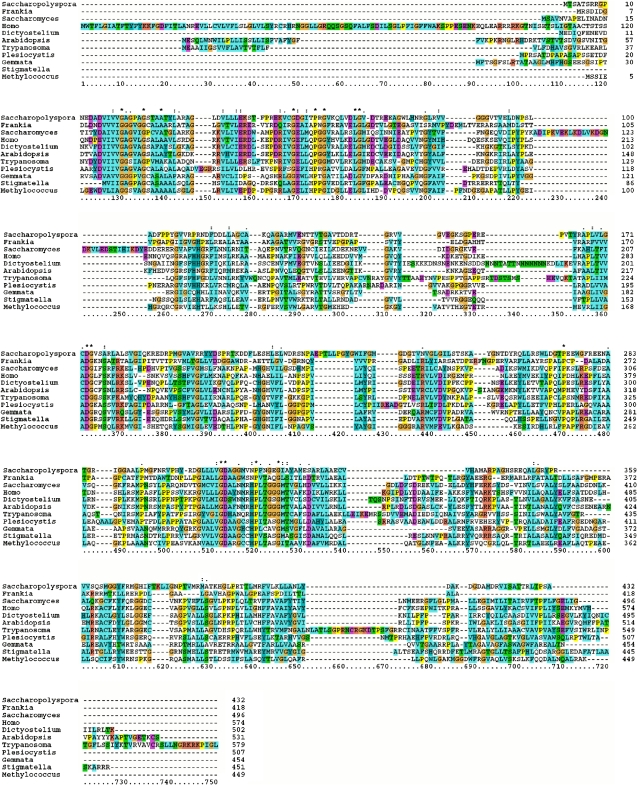
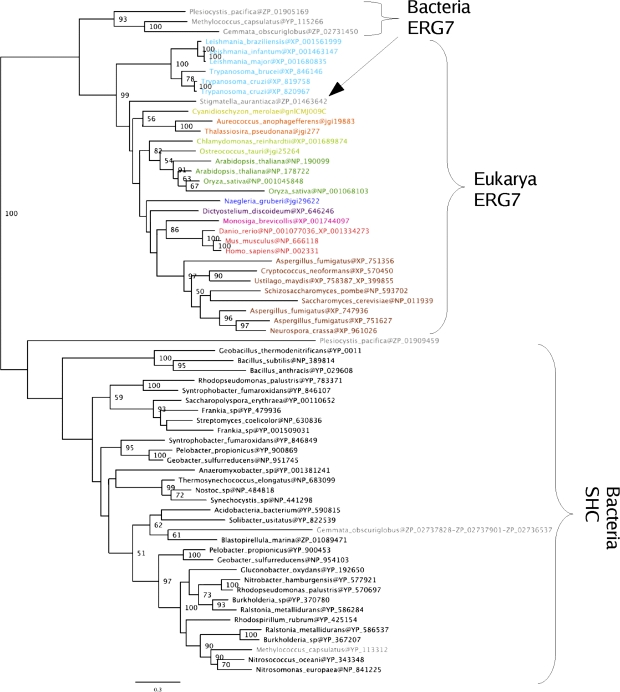
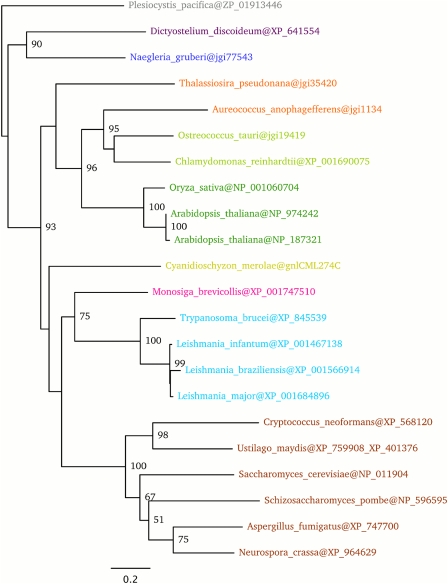
The availability of complete genomes from a wide sampling of eukaryotic diversity has allowed the application of phylogenomics approaches to study the origin and evolution of unique eukaryotic cellular structures, but these are still poorly applied to study unique eukaryotic metabolic pathways. Sterols are a good example because they are an essential feature of eukaryotic membranes. The sterol pathway has been well dissected in vertebrates, fungi, and land plants. However, although different types of sterols have been identified in other eukaryotic lineages, their pathways have not been fully characterized. We have carried out an extensive analysis of the taxonomic distribution and phylogeny of the enzymes of the sterol pathway in a large sampling of eukaryotic lineages. This allowed us to tentatively indicate features of the sterol pathway in organisms where this has not been characterized and to point out a number of steps for which yet-to-discover enzymes may be at work. We also inferred that the last eukaryotic common ancestor already harbored a large panel of enzymes for sterol synthesis and that subsequent evolution over the eukaryotic tree occurred by tinkering, mainly by gene losses. We highlight a high capacity of sterol synthesis in the myxobacterium Plesiocystis pacifica, and we support the hypothesis that the few bacteria that harbor homologs of the sterol pathway have likely acquired these via horizontal gene transfer from eukaryotes. Finally, we propose a potential candidate for the elusive enzyme performing C-3 ketoreduction (ERG27 equivalent) in land plants and probably in other eukaryotic phyla.
Keywords: eukaryotes; evolution; phylogenomics; sterols.
Figures


References
-
- Anderson RGW, Jacobson K. Cell biology—a role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science. 2002;296:1821–1825. - PubMed
-
- Andersson JO. Horizontal gene transfer between microbial Eukaryotes. In: Gogarten MB, Gogarten JP, Olendzenski L, editors. Horizontal gene transfer: genomes in flux. Totowa (NJ): Humana Press Inc; 2009. pp. 473–487.
-
- Bloch K. Cholesterol: evolution of structure and function. In: Vance DE, Vance J, editors. Biochemistry of lipids, lipoproteins and membranes. Amsterdam: Elsevier; 1991. pp. 363–381.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
