

Mobile regulatory cassettes mediate modular shuffling in T4-type phage genomes
- PMID: 20333230
- PMCID: PMC2839356
- DOI: 10.1093/gbe/evq006
Mobile regulatory cassettes mediate modular shuffling in T4-type phage genomes
Abstract
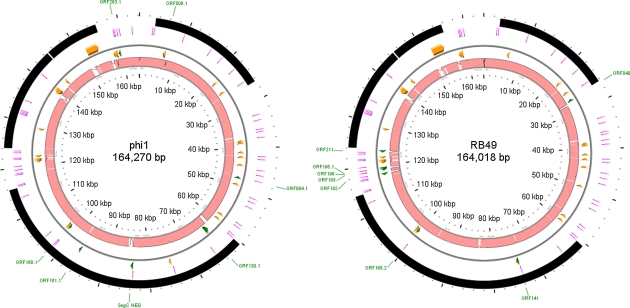
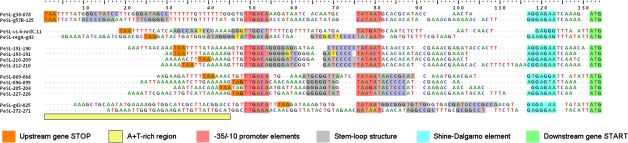
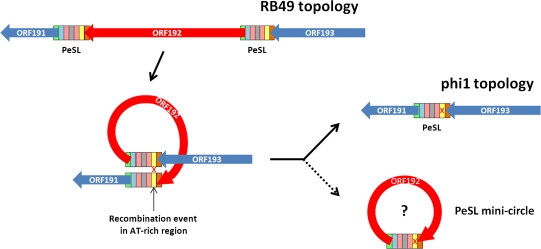
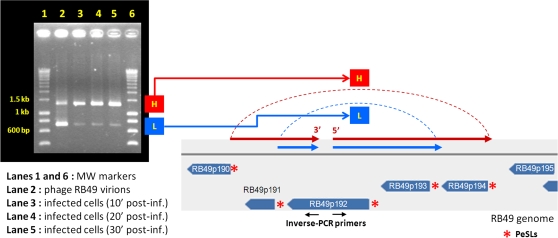
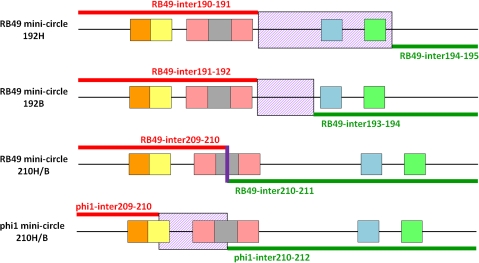
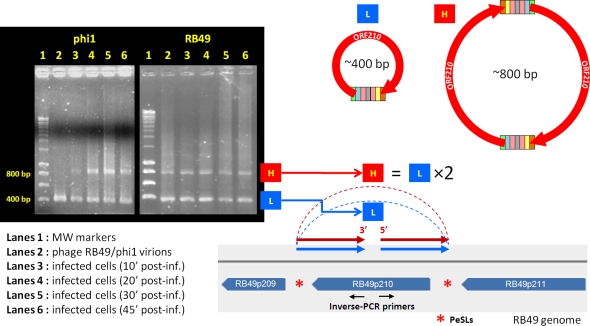
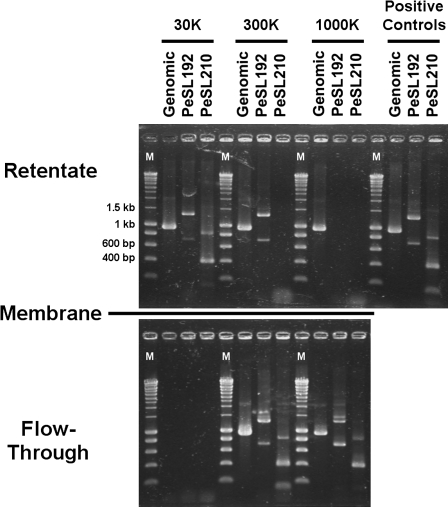
Coliphage phi1, which was isolated for phage therapy in the Republic of Georgia, is closely related to the T-like myovirus RB49. The approximately 275 open reading frames encoded by each phage have an average level of amino acid identity of 95.8%. RB49 lacks 7 phi1 genes while 10 phi1 genes are missing from RB49. Most of these unique genes encode functions without known homologs. Many of the insertion, deletion, and replacement events that distinguish the two phages are in the hyperplastic regions (HPRs) of their genomes. The HPRs are rich in both nonessential genes and small regulatory cassettes (promoter(early) stem-loops [PeSLs]) composed of strong sigma(70)-like promoters and stem-loop structures, which are effective transcription terminators. Modular shuffling mediated by recombination between PeSLs has caused much of the sequence divergence between RB49 and phi1. We show that exchanges between nearby PeSLs can also create small circular DNAs that are apparently encapsidated by the virus. Such PeSL "mini-circles" may be important vectors for horizontal gene transfer.
Keywords: T4-like phage; genome evolution; modular shuffling; regulatory cassette.
Figures
References
-
- Ackermann HW, Krisch HM. A catalogue of T4-type bacteriophages. Arch Virol. 1997;142:2329–2345. - PubMed
-
- Ausubel FM, et al. Short protocols in molecular biology. New York: J. Wiley and Sons; 1992.
-
- Botstein D. A theory of modular evolution for bacteriophages. Ann N Y Acad Sci. 1980;354:484–491. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
