

The Salmonella type III secretion effector, salmonella leucine-rich repeat protein (SlrP), targets the human chaperone ERdj3
- PMID: 20335166
- PMCID: PMC2871503
- DOI: 10.1074/jbc.M110.100669
The Salmonella type III secretion effector, salmonella leucine-rich repeat protein (SlrP), targets the human chaperone ERdj3
Abstract
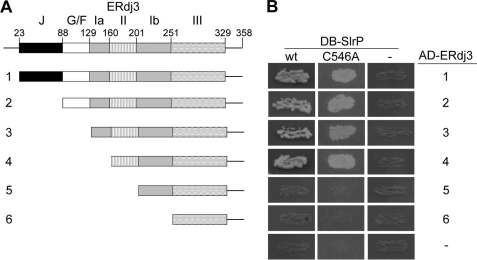
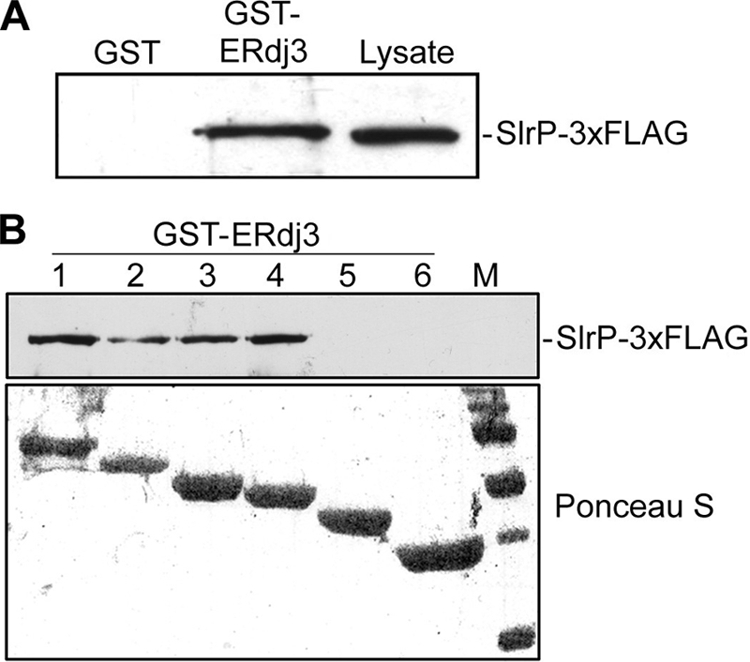
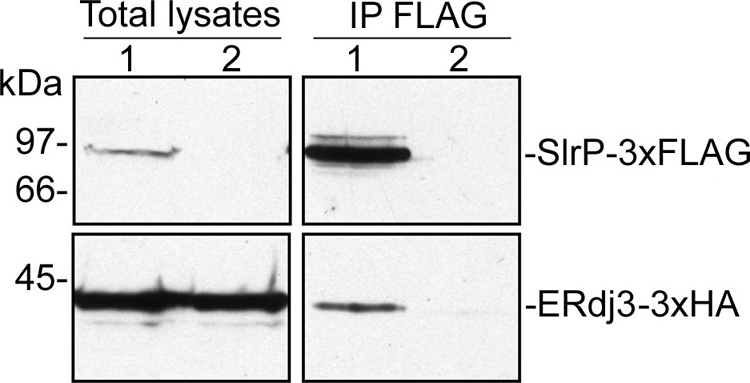
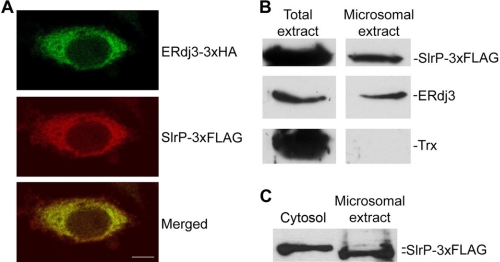
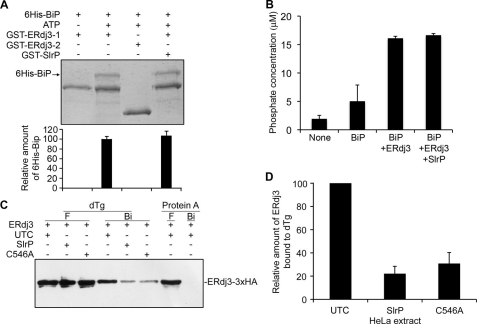
Effectors of the type III secretion systems (T3SS) are key elements in the interaction between many Gram-negative pathogens and their hosts. SlrP is an effector that is translocated into the eukaryotic host cell through the two virulence-associated T3SS of Salmonella enterica. We found previously that this effector is an E3 ubiquitin ligase for mammalian thioredoxin. Here, we identified ERdj3, an endoplasmic reticulum lumenal chaperone of the Hsp40/DnaJ family, as a new target for SlrP. Experiments with truncated forms of ERdj3 showed that domain II was essential for the interaction with SlrP. Confocal microscopy and subcellular fractionation demonstrated that, in transfected HeLa cells, SlrP was partially located in the endoplasmic reticulum. The presence of SlrP interfered with the binding of ERdj3 to a denatured substrate. Taken together, these data suggest that the role of SlrP in the interaction between Salmonella and the host cell is exerted through the modulation of the function of two independent targets: thioredoxin in the cytosol, and ERdj3 in the endoplasmic reticulum.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
