

Mathematical modeling of ultradeep sequencing data reveals that acute CD8+ T-lymphocyte responses exert strong selective pressure in simian immunodeficiency virus-infected macaques but still fail to clear founder epitope sequences
- PMID: 20335256
- PMCID: PMC2876615
- DOI: 10.1128/JVI.00117-10
Mathematical modeling of ultradeep sequencing data reveals that acute CD8+ T-lymphocyte responses exert strong selective pressure in simian immunodeficiency virus-infected macaques but still fail to clear founder epitope sequences
Abstract
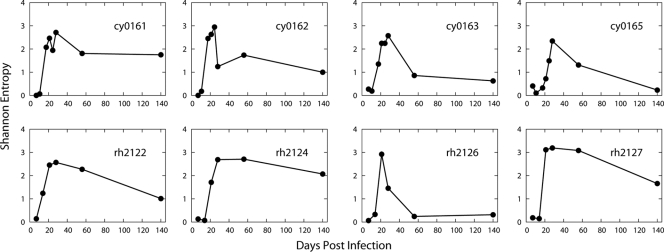
The prominent role of antiviral cytotoxic CD8(+) T-lymphocytes (CD8-TL) in containing the acute viremia of human and simian immunodeficiency viruses (HIV-1 and SIV) has rationalized the development of T-cell-based vaccines. However, the presence of escape mutations in the acute stage of infection has raised a concern that accelerated escape from vaccine-induced CD8-TL responses might undermine vaccine efficacy. We reanalyzed previously published data of 101,822 viral genomes of three CD8-TL epitopes, Nef(103-111)RM9 (RM9), Tat(28-35)SL8 (SL8), and Gag(181-189)CM9 (CM9), sampled by ultradeep pyrosequencing from eight macaques. Multiple epitope variants appeared during the resolution of acute viremia, followed by the predominance of a single mutant epitope. By fitting a mathematical model, we estimated the first acute escape rate as 0.36 day(-1) within escape-prone epitopes, RM9 and SL8, and the chronic escape rate as 0.014 day(-1) within the CM9 epitope. Our estimate of SIV acute escape rates was found to be comparable to very early HIV-1 escape rates. The timing of the first escape was more highly correlated with the timing of the peak CD8-TL response than with the magnitude of the CD8-TL response. The transmitted epitope decayed more than 400 times faster during the acute viral decline stage than predicted by a neutral evolution model. However, the founder epitope persisted as a minor population even at the viral set point; in contrast, the majority of acute escape epitopes were completely cleared. Our results suggest that a reservoir of SIV infection is preferentially formed by virus with the transmitted epitope.
Figures






References
-
- Abrahams, M. R., J. A. Anderson, E. E. Giorgi, C. Seoighe, K. Mlisana, L. H. Ping, G. S. Athreya, F. K. Treurnicht, B. F. Keele, N. Wood, J. F. Salazar-Gonzalez, T. Bhattacharya, H. Chu, I. Hoffman, S. Galvin, C. Mapanje, P. Kazembe, R. Thebus, S. Fiscus, W. Hide, M. S. Cohen, S. A. Karim, B. F. Haynes, G. M. Shaw, B. H. Hahn, B. T. Korber, R. Swanstrom, and C. Williamson. 2009. Quantitating the multiplicity of infection with human immunodeficiency virus type 1 subtype C reveals a non-Poisson distribution of transmitted variants. J. Virol. 83:3556-3567. - PMC - PubMed
-
- Addo, M. M., R. Draenert, A. Rathod, C. L. Verrill, B. T. Davis, R. T. Gandhi, G. K. Robbins, N. O. Basgoz, D. R. Stone, D. E. Cohen, M. N. Johnston, T. Flynn, A. G. Wurcel, E. S. Rosenberg, M. Altfeld, and B. D. Walker. 2007. Fully differentiated HIV-1 specific CD8+ T effector cells are more frequently detectable in controlled than in progressive HIV-1 infection. PLoS One 2:e321. - PMC - PubMed
-
- Addo, M. M., X. G. Yu, A. Rathod, D. Cohen, R. L. Eldridge, D. Strick, M. N. Johnston, C. Corcoran, A. G. Wurcel, C. A. Fitzpatrick, M. E. Feeney, W. R. Rodriguez, N. Basgoz, R. Draenert, D. R. Stone, C. Brander, P. J. Goulder, E. S. Rosenberg, M. Altfeld, and B. D. Walker. 2003. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J. Virol. 77:2081-2092. - PMC - PubMed
-
- Agresti, A. 2002. Categorical data analysis, 2nd ed. John Wiley & Sons, Hoboken, NJ.
-
- Allen, T. M., D. H. O'Connor, P. Jing, J. L. Dzuris, B. R. Mothe, T. U. Vogel, E. Dunphy, M. E. Liebl, C. Emerson, N. Wilson, K. J. Kunstman, X. Wang, D. B. Allison, A. L. Hughes, R. C. Desrosiers, J. D. Altman, S. M. Wolinsky, A. Sette, and D. I. Watkins. 2000. Tat-specific cytotoxic T lymphocytes select for SIV escape variants during resolution of primary viraemia. Nature 407:386-390. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
