

Role of complex carbohydrates in human immunodeficiency virus type 1 infection and resistance to antibody neutralization
- PMID: 20335257
- PMCID: PMC2876609
- DOI: 10.1128/JVI.00105-10
Role of complex carbohydrates in human immunodeficiency virus type 1 infection and resistance to antibody neutralization
Abstract
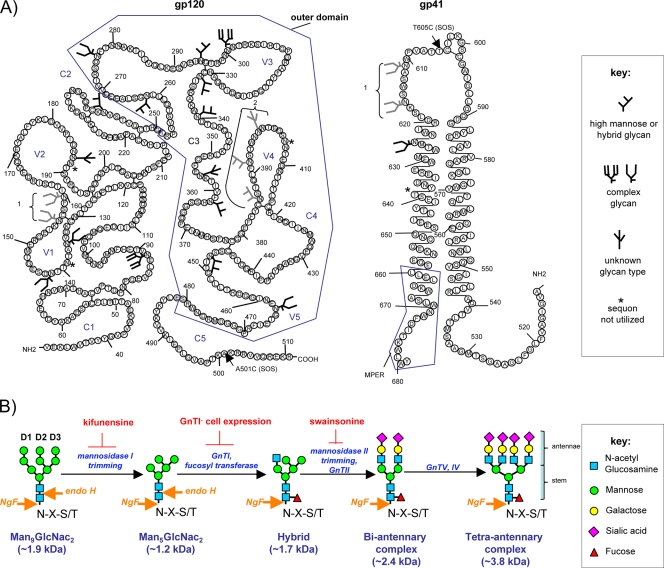
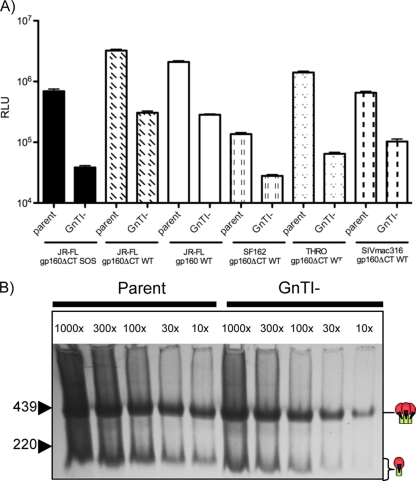
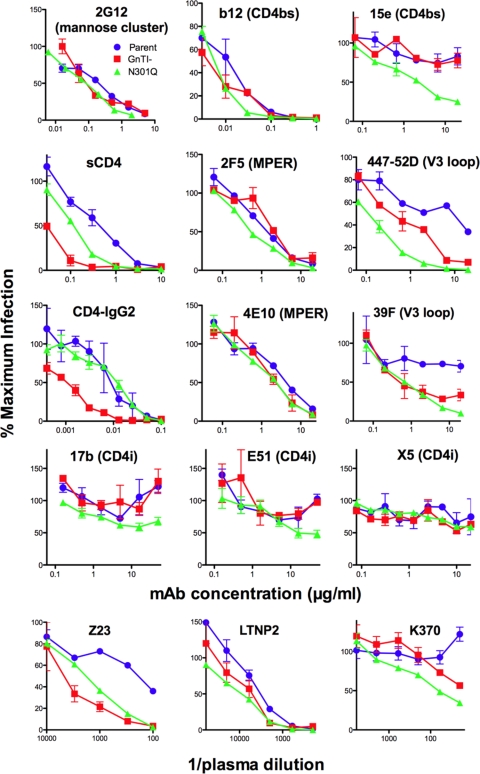
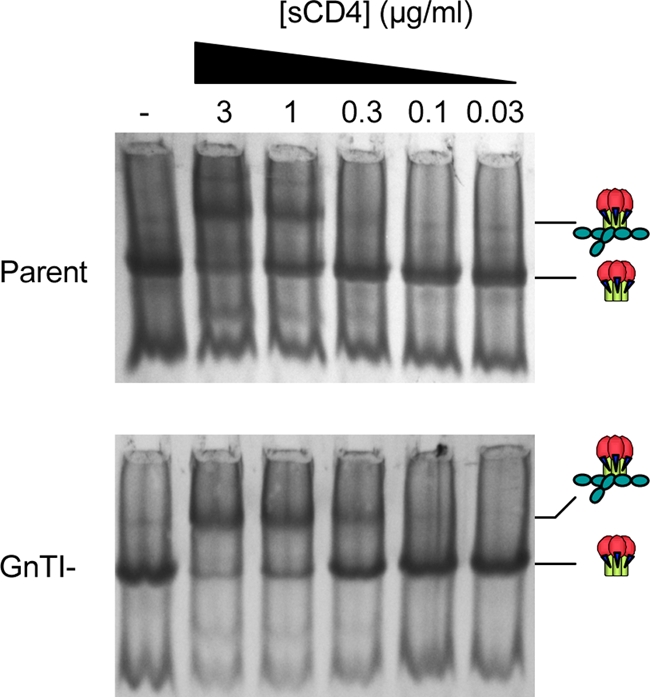
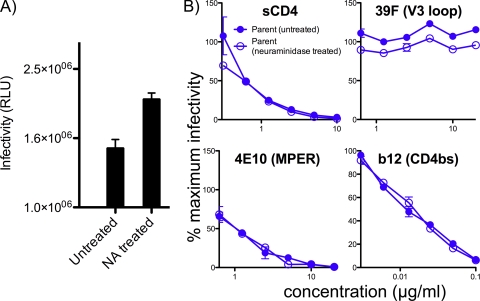
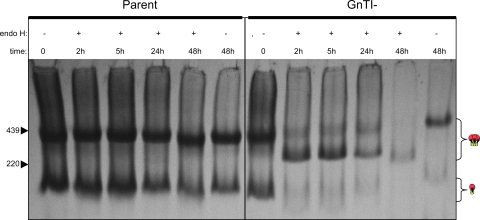
Complex N-glycans flank the receptor binding sites of the outer domain of HIV-1 gp120, ostensibly forming a protective "fence" against antibodies. Here, we investigated the effects of rebuilding this fence with smaller glycoforms by expressing HIV-1 pseudovirions from a primary isolate in a human cell line lacking N-acetylglucosamine transferase I (GnTI), the enzyme that initiates the conversion of oligomannose N-glycans into complex N-glycans. Thus, complex glycans, including those that surround the receptor binding sites, are replaced by fully trimmed oligomannose stumps. Conversely, the untrimmed oligomannoses of the silent domain of gp120 are likely to remain unchanged. For comparison, we produced a mutant virus lacking a complex N-glycan of the V3 loop (N301Q). Both variants exhibited increased sensitivities to V3 loop-specific monoclonal antibodies (MAbs) and soluble CD4. The N301Q virus was also sensitive to "nonneutralizing" MAbs targeting the primary and secondary receptor binding sites. Endoglycosidase H treatment resulted in the removal of outer domain glycans from the GnTI- but not the parent Env trimers, and this was associated with a rapid and complete loss in infectivity. Nevertheless, the glycan-depleted trimers could still bind to soluble receptor and coreceptor analogs, suggesting a block in post-receptor binding conformational changes necessary for fusion. Collectively, our data show that the antennae of complex N-glycans serve to protect the V3 loop and CD4 binding site, while N-glycan stems regulate native trimer conformation, such that their removal can lead to global changes in neutralization sensitivity and, in extreme cases, an inability to complete the conformational rearrangements necessary for infection.
Figures

Similar articles
-
Conserved Role of an N-Linked Glycan on the Surface Antigen of Human Immunodeficiency Virus Type 1 Modulating Virus Sensitivity to Broadly Neutralizing Antibodies against the Receptor and Coreceptor Binding Sites.J Virol. 2015 Oct 28;90(2):829-41. doi: 10.1128/JVI.02321-15. Print 2016 Jan 15. J Virol. 2015. PMID: 26512079 Free PMC article.
-
N-linked glycosylation of the V3 loop and the immunologically silent face of gp120 protects human immunodeficiency virus type 1 SF162 from neutralization by anti-gp120 and anti-gp41 antibodies.J Virol. 2004 Apr;78(7):3279-95. doi: 10.1128/jvi.78.7.3279-3295.2004. J Virol. 2004. PMID: 15016849 Free PMC article.
-
Structure-based, targeted deglycosylation of HIV-1 gp120 and effects on neutralization sensitivity and antibody recognition.Virology. 2003 Sep 1;313(2):387-400. doi: 10.1016/s0042-6822(03)00294-0. Virology. 2003. PMID: 12954207
-
Antigenicity and Immunogenicity of HIV-1 Envelope Trimers Complexed to a Small-Molecule Viral Entry Inhibitor.J Virol. 2020 Oct 14;94(21):e00958-20. doi: 10.1128/JVI.00958-20. Print 2020 Oct 14. J Virol. 2020. PMID: 32817216 Free PMC article.
-
Changes in Structure and Antigenicity of HIV-1 Env Trimers Resulting from Removal of a Conserved CD4 Binding Site-Proximal Glycan.J Virol. 2016 Sep 29;90(20):9224-36. doi: 10.1128/JVI.01116-16. Print 2016 Oct 15. J Virol. 2016. PMID: 27489265 Free PMC article.
Cited by
-
Functional and Structural Characterization of Human V3-Specific Monoclonal Antibody 2424 with Neutralizing Activity against HIV-1 JRFL.J Virol. 2015 Sep;89(17):9090-102. doi: 10.1128/JVI.01280-15. Epub 2015 Jun 24. J Virol. 2015. PMID: 26109728 Free PMC article.
-
Asymmetric recognition of the HIV-1 trimer by broadly neutralizing antibody PG9.Proc Natl Acad Sci U S A. 2013 Mar 12;110(11):4351-6. doi: 10.1073/pnas.1217537110. Epub 2013 Feb 20. Proc Natl Acad Sci U S A. 2013. PMID: 23426631 Free PMC article.
-
Single genome amplification and standard bulk PCR yield HIV-1 envelope products with similar genotypic and phenotypic characteristics.J Virol Methods. 2015 Mar;214:46-53. doi: 10.1016/j.jviromet.2015.01.006. Epub 2015 Feb 11. J Virol Methods. 2015. PMID: 25681527 Free PMC article.
-
Elicitation of broadly reactive antibodies against glycan-modulated neutralizing V3 epitopes of HIV-1 by immune complex vaccines.Vaccine. 2013 Nov 4;31(46):5413-21. doi: 10.1016/j.vaccine.2013.09.010. Epub 2013 Sep 16. Vaccine. 2013. PMID: 24051158 Free PMC article.
-
A human antibody to the CD4 binding site of gp120 capable of highly potent but sporadic cross clade neutralization of primary HIV-1.PLoS One. 2013 Aug 26;8(8):e72054. doi: 10.1371/journal.pone.0072054. eCollection 2013. PLoS One. 2013. PMID: 23991039 Free PMC article.
References
-
- Allan, J. S., J. Strauss, and D. W. Buck. 1990. Enhancement of SIV infection with soluble receptor molecules. Science 247:1084-1088. - PubMed
-
- Astronomo, R. D., H. K. Lee, C. N. Scanlan, R. Pantophlet, C. Y. Huang, I. A. Wilson, O. Blixt, R. A. Dwek, C. H. Wong, and D. R. Burton. 2008. A glycoconjugate antigen based on the recognition motif of a broadly neutralizing human immunodeficiency virus antibody, 2G12, is immunogenic but elicits antibodies unable to bind to the self glycans of gp120. J. Virol. 82:6359-6368. - PMC - PubMed
-
- Back, N. K., L. Smit, J. J. De Jong, W. Keulen, M. Schutten, J. Goudsmit, and M. Tersmette. 1994. An N-glycan within the human immunodeficiency virus type 1 gp120 V3 loop affects virus neutralization. Virology 199:431-438. - PubMed
-
- Barbas, C. F., III, T. A. Collet, W. Amberg, P. Roben, J. M. Binley, D. Hoekstra, D. Cababa, T. M. Jones, R. A. Williamson, G. R. Pilkington, et al. 1993. Molecular profile of an antibody response to HIV-1 as probed by combinatorial libraries. J. Mol. Biol. 230:812-823. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
