

Role of annexin A2 in the production of infectious hepatitis C virus particles
- PMID: 20335258
- PMCID: PMC2876593
- DOI: 10.1128/JVI.02343-09
Role of annexin A2 in the production of infectious hepatitis C virus particles
Abstract
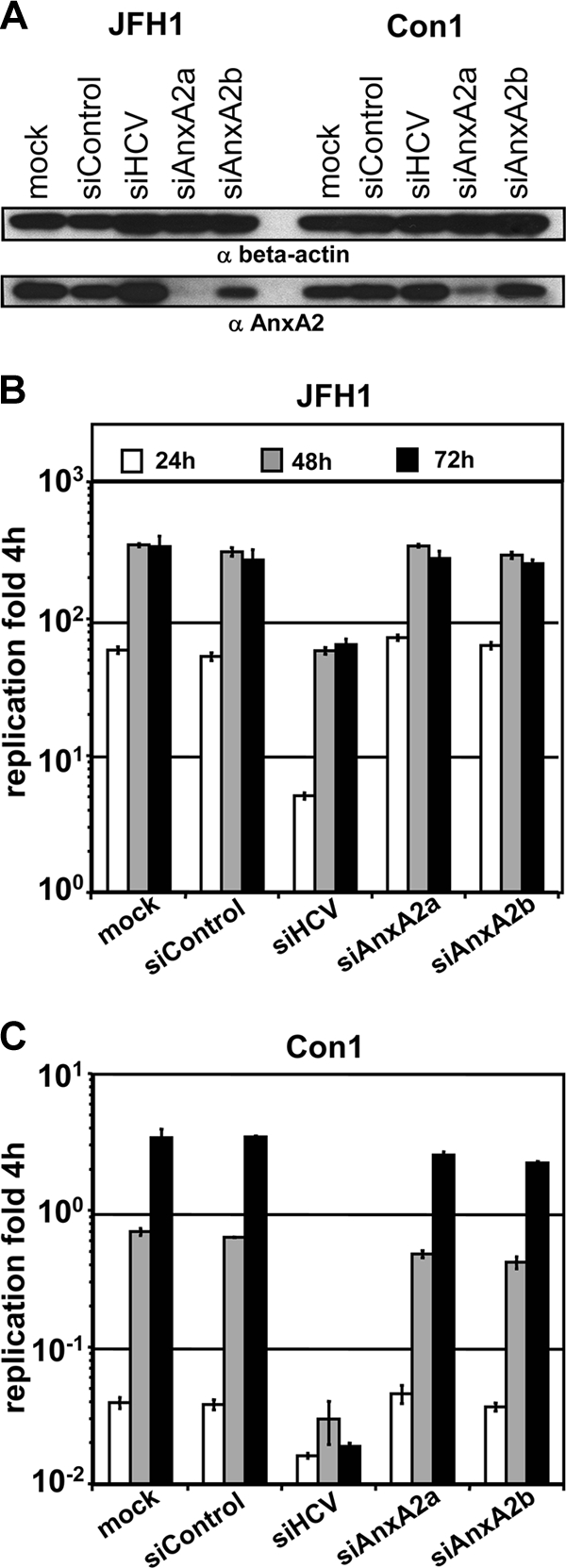
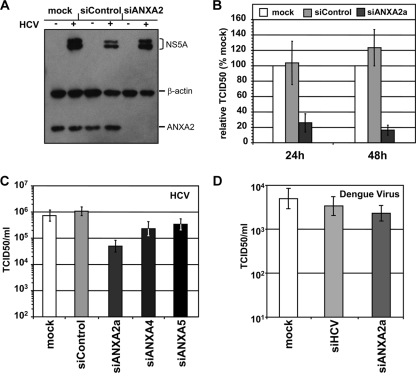
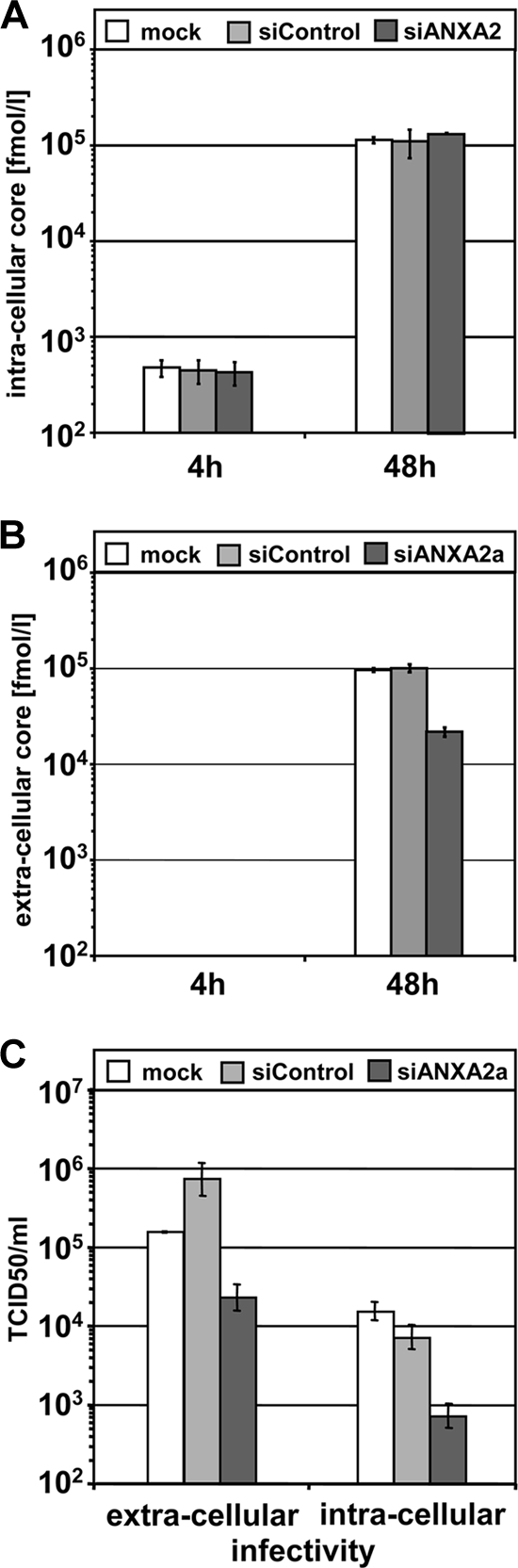
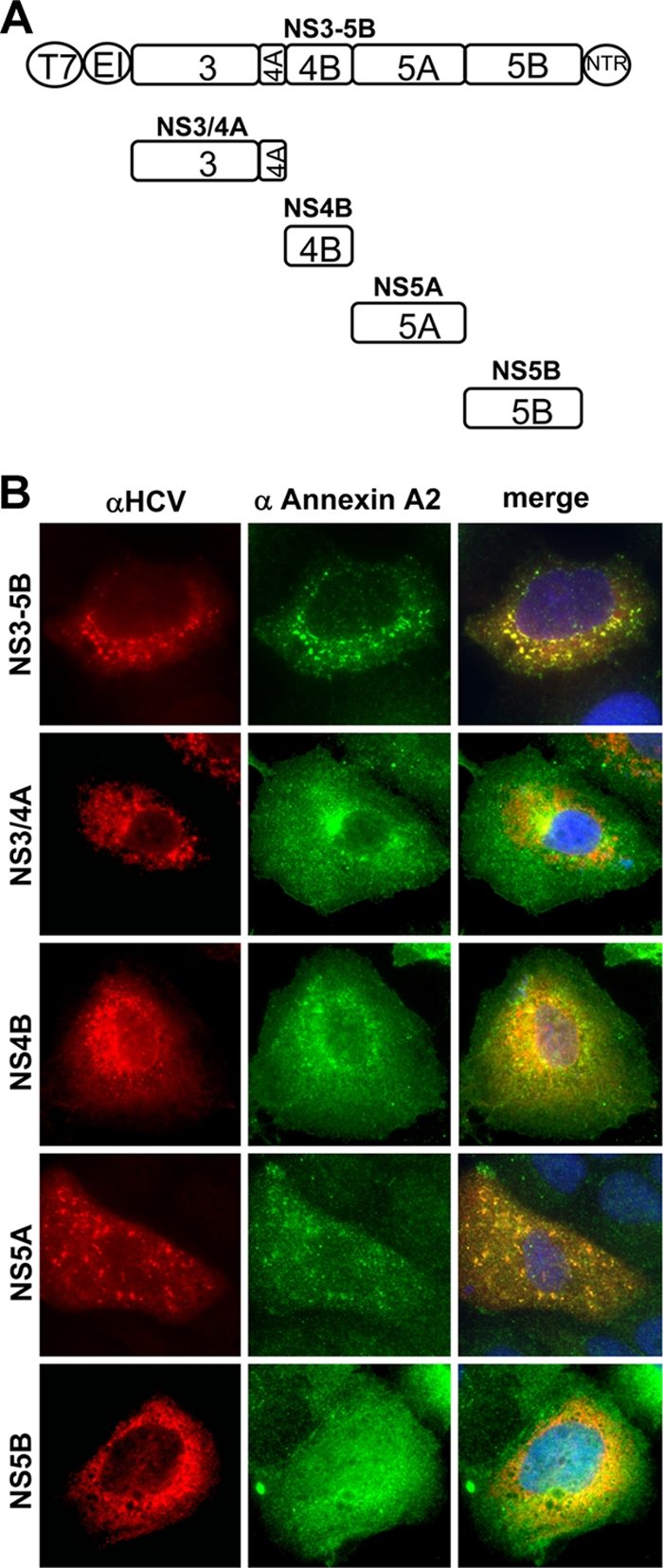
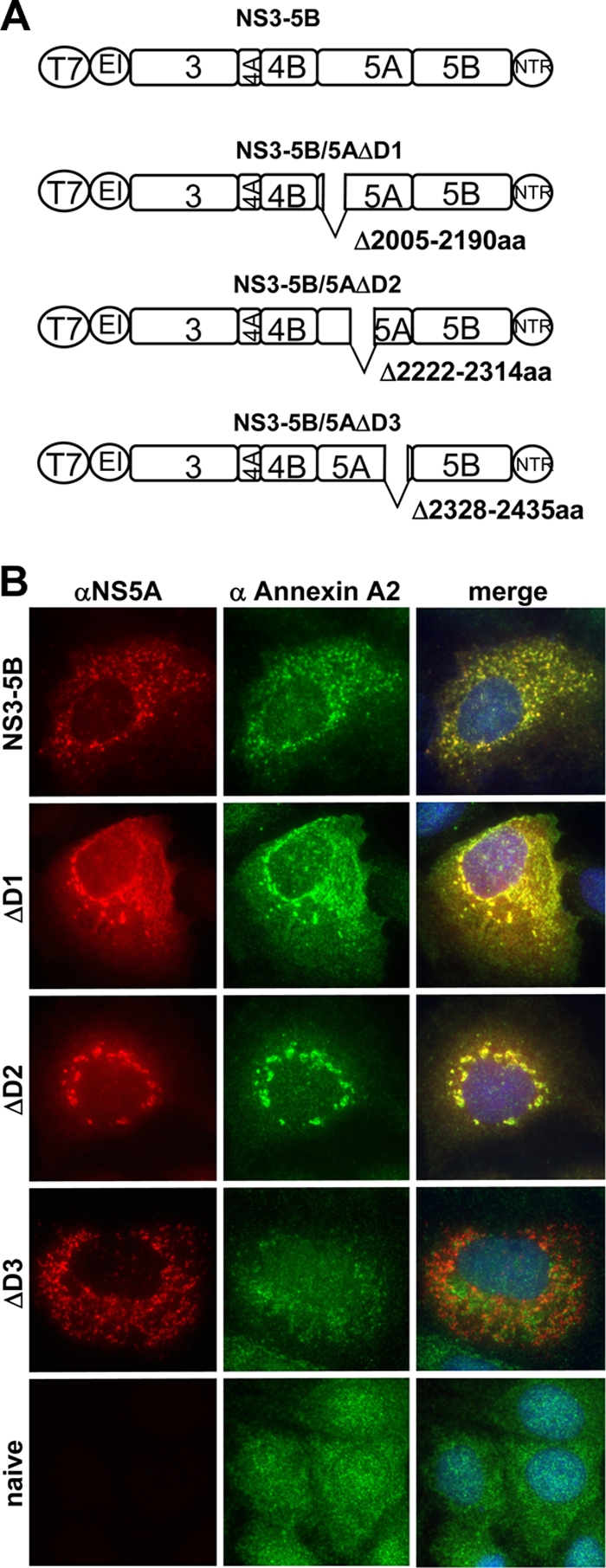
Hepatitis C virus (HCV) is an important human pathogen affecting 170 million chronically infected individuals. In search for cellular proteins involved in HCV replication, we have developed a purification strategy for viral replication complexes and identified annexin A2 (ANXA2) as an associated host factor. ANXA2 colocalized with viral nonstructural proteins in cells harboring genotype 1 or 2 replicons as well as in infected cells. In contrast, we found no obvious colocalization of ANXA2 with replication sites of other positive-strand RNA viruses. The silencing of ANXA2 expression showed no effect on viral RNA replication but resulted in a significant reduction of extra- and intracellular virus titers. Therefore, it seems likely that ANXA2 plays a role in HCV assembly rather than in genome replication or virion release. Colocalization studies with individually expressed HCV nonstructural proteins indicated that NS5A specifically recruits ANXA2, probably by an indirect mechanism. By the deletion of individual NS5A subdomains, we identified domain III (DIII) as being responsible for ANXA2 recruitment. These data identify ANXA2 as a novel host factor contributing, with NS5A, to the formation of infectious HCV particles.
Figures
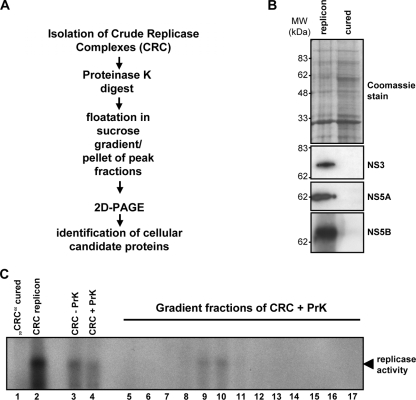
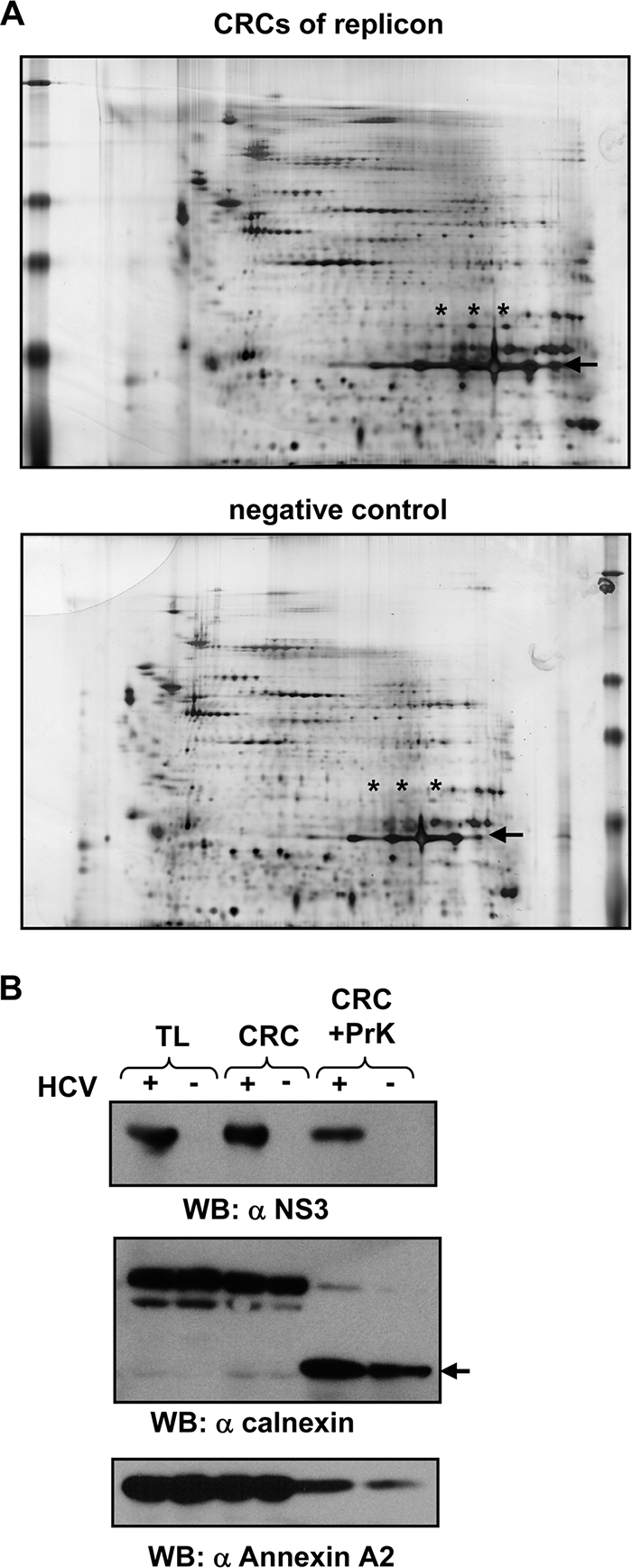
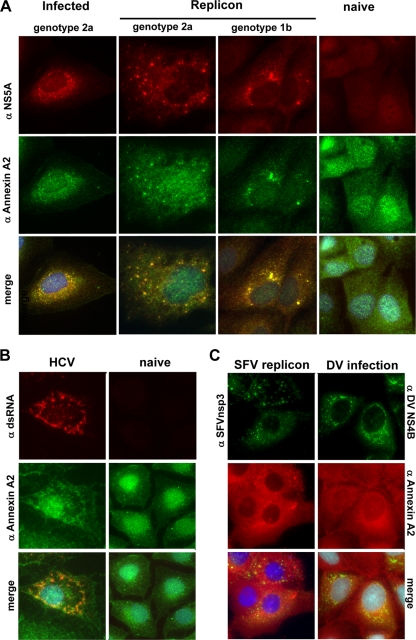
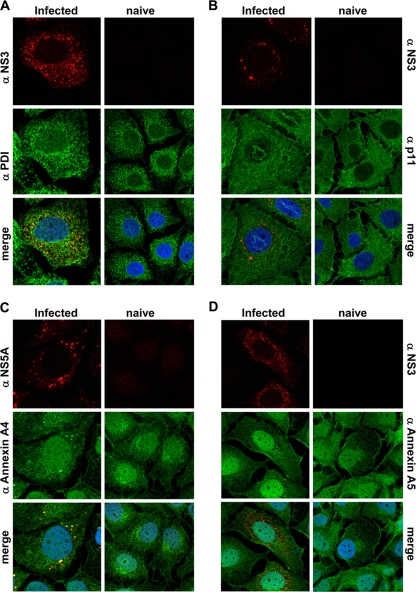
References
-
- Bartenschlager, R., M. Frese, and T. Pietschmann. 2004. Novel insights into hepatitis C virus replication and persistence. Adv. Virus Res. 63:71-180. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
