

Deregulated copper transport affects Arabidopsis development especially in the absence of environmental cycles
- PMID: 20335405
- PMCID: PMC2862424
- DOI: 10.1104/pp.110.153676
Deregulated copper transport affects Arabidopsis development especially in the absence of environmental cycles
Abstract
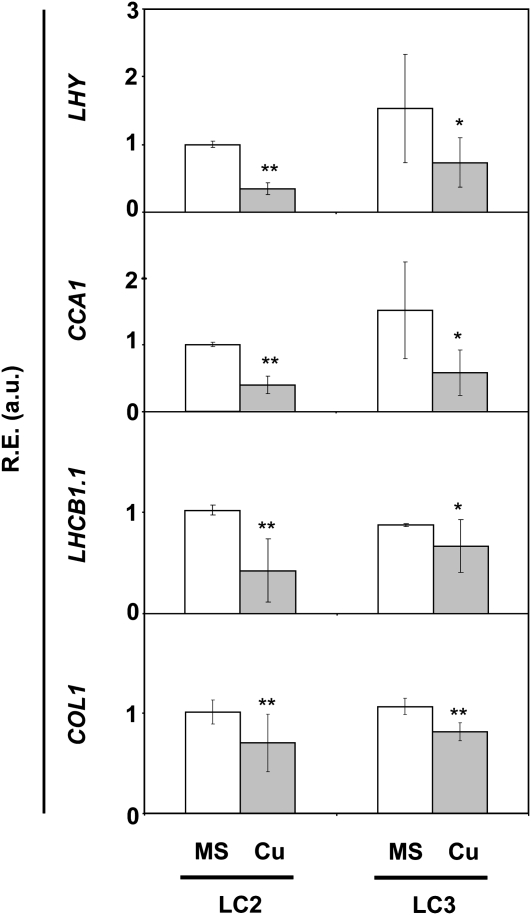
Copper is an essential cofactor for key processes in plants, but it exerts harmful effects when in excess. Previous work has shown that the Arabidopsis (Arabidopsis thaliana) COPT1 high-affinity copper transport protein participates in copper uptake through plant root tips. Here, we show that COPT1 protein localizes to the plasma membrane of Arabidopsis cells and the phenotypic effects of transgenic plants overexpressing either COPT1 or COPT3, the latter being another high-affinity copper transport protein family member. Both transgenic lines exhibit increased endogenous copper levels and are sensitive to the copper in the growth medium. Additional phenotypes include decreased hypocotyl growth in red light and differentially affected flowering times depending on the photoperiod. Furthermore, in the absence of environmental cycles, such as light and temperature, the survival of plants overexpressing COPT1 or COPT3 is compromised. Consistent with altered circadian rhythms, the expression of the nuclear circadian clock genes CIRCADIAN CLOCK-ASSOCIATED1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) is substantially reduced in either COPT1- or COPT3-overexpressing plants. Copper affects the amplitude and the phase, but not the period, of the CCA1 and LHY oscillations in wild-type plants. Copper also drives a reduction in the expression of circadian clock output genes. These results reveal that the spatiotemporal control of copper transport is a key aspect of metal homeostasis that is required for Arabidopsis fitness, especially in the absence of environmental cues.
Figures









References
-
- Andrés-Colás N, Sancenon V, Rodriguez-Navarro S, Mayo S, Thiele DJ, Ecker JR, Puig S, Peñarrubia L. (2006) The Arabidopsis heavy metal P-type ATPase HMA5 interacts with metallochaperones and functions in copper detoxification of roots. Plant J 45: 225–236 - PubMed
-
- Bellemare DR, Shaner L, Morano KA, Beaudoin J, Langlois R, Labbe S. (2002) Ctr6, a vacuolar membrane copper transporter in Schizosaccharomyces pombe. J Biol Chem 277: 46676–46686 - PubMed
-
- Bertinato J, Swist E, Plouffe LJ, Brooks SP, L'Abbe MR. (2008) Ctr2 is partially localized to the plasma membrane and stimulates copper uptake in COS-7 cells. Biochem J 409: 731–740 - PubMed
-
- Buege JA, Aust SD. (1978) Microsomal lipid peroxidation. Methods Enzymol 52: 302–310 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
