

MLT-10 defines a family of DUF644 and proline-rich repeat proteins involved in the molting cycle of Caenorhabditis elegans
- PMID: 20335506
- PMCID: PMC2869372
- DOI: 10.1091/mbc.e08-07-0708
MLT-10 defines a family of DUF644 and proline-rich repeat proteins involved in the molting cycle of Caenorhabditis elegans
Abstract
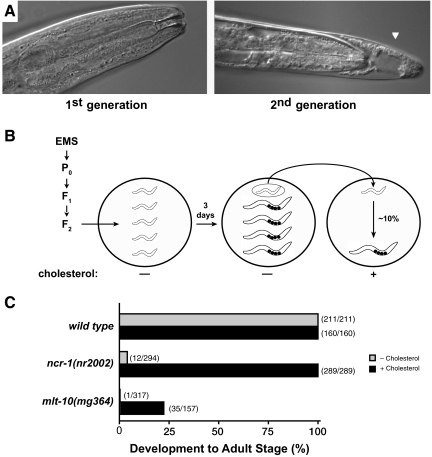
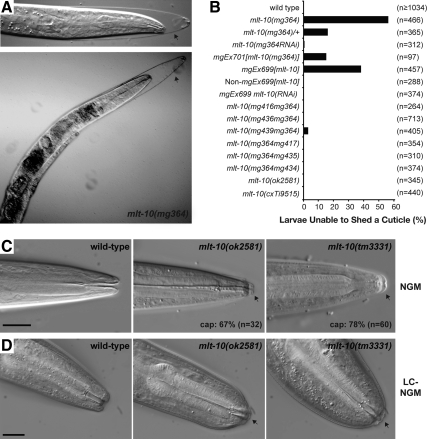
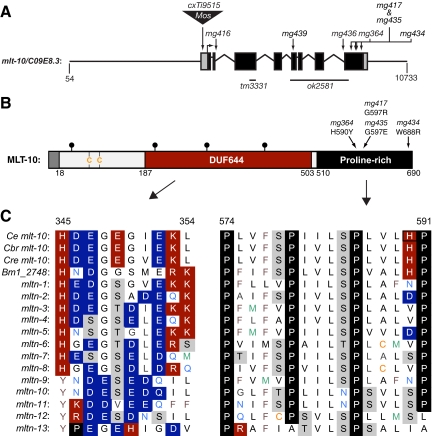
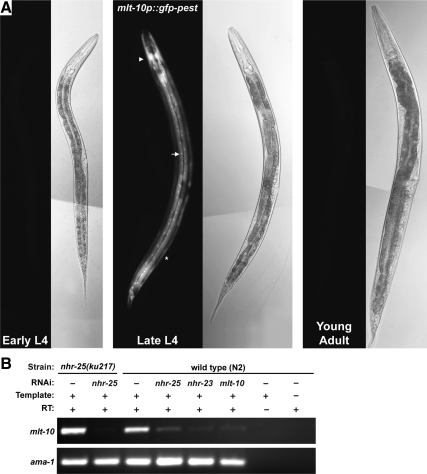
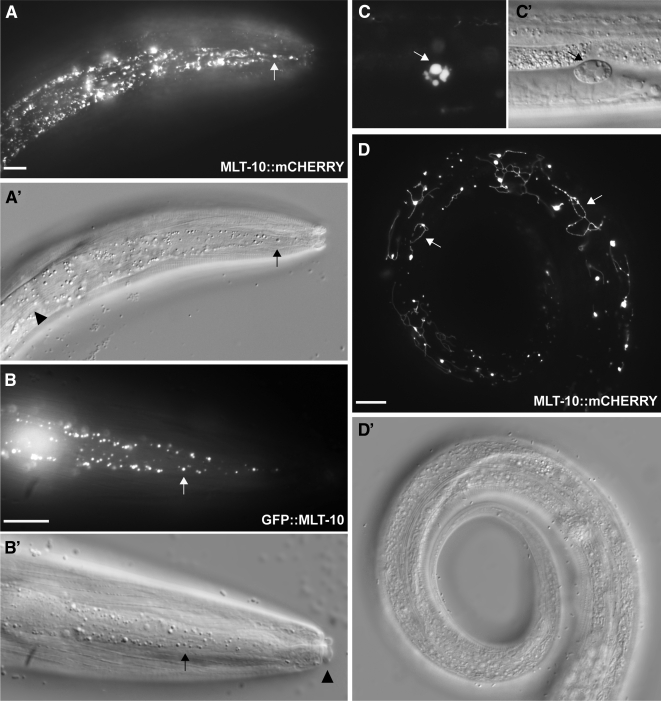
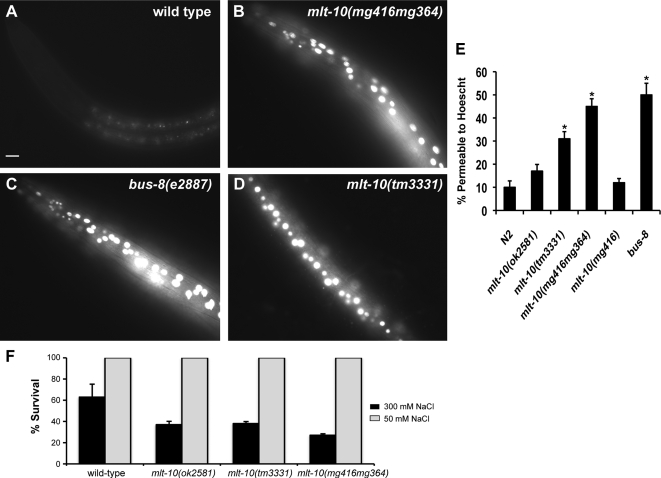
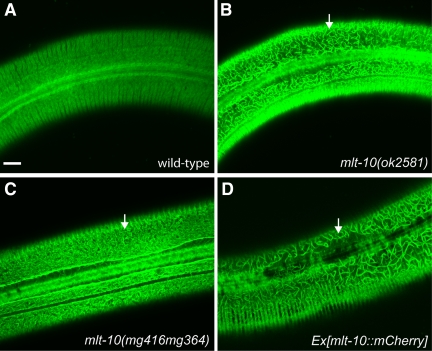
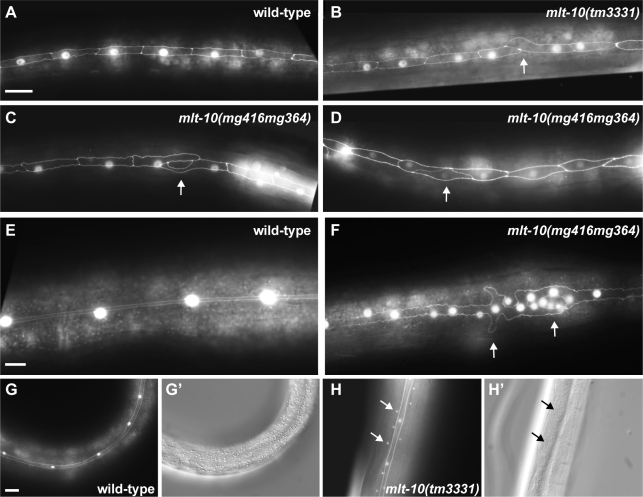
The molting cycle of nematodes involves the periodic synthesis and removal of a collagen-rich exoskeleton, but the underlying molecular mechanisms are not well understood. Here, we describe the mlt-10 gene of Caenorhabditis elegans, which emerged from a genetic screen for molting-defective mutants sensitized by low cholesterol. MLT-10 defines a large family of nematode-specific proteins comprised of DUF644 and tandem P-X(2)-L-(S/T)-P repeats. Conserved nuclear hormone receptors promote expression of the mlt-10 gene in the hypodermis whenever the exoskeleton is remade. Further, a MLT-10::mCherry fusion protein is released from the hypodermis to the surrounding matrices and fluids during molting. The fusion protein is also detected in strands near the surface of animals. Both loss-of-function and gain-of-function mutations of mlt-10 impede the removal of old cuticles. However, the substitution mutation mlt-10(mg364), which disrupts the proline-rich repeats, causes the most severe phenotype. Mutations of mlt-10 are also associated with abnormalities in the exoskeleton and improper development of the epidermis. Thus, mlt-10 encodes a secreted protein involved in three distinct but interconnected aspects of the molting cycle. We propose that the molting cycle of C. elegans involves the dynamic assembly and disassembly of MLT-10 and possibly the paralogs of MLT-10.
Figures

Similar articles
-
Conserved Ankyrin Repeat Proteins and Their NIMA Kinase Partners Regulate Extracellular Matrix Remodeling and Intracellular Trafficking in Caenorhabditis elegans.Genetics. 2017 Jan;205(1):273-293. doi: 10.1534/genetics.116.194464. Epub 2016 Oct 31. Genetics. 2017. PMID: 27799278 Free PMC article.
-
Actin organization and endocytic trafficking are controlled by a network linking NIMA-related kinases to the CDC-42-SID-3/ACK1 pathway.PLoS Genet. 2018 Apr 2;14(4):e1007313. doi: 10.1371/journal.pgen.1007313. eCollection 2018 Apr. PLoS Genet. 2018. PMID: 29608564 Free PMC article.
-
The mir-84 and let-7 paralogous microRNA genes of Caenorhabditis elegans direct the cessation of molting via the conserved nuclear hormone receptors NHR-23 and NHR-25.Development. 2006 Dec;133(23):4631-41. doi: 10.1242/dev.02655. Epub 2006 Oct 25. Development. 2006. PMID: 17065234
-
Cuticle collagen genes. Expression in Caenorhabditis elegans.Trends Genet. 2000 Jan;16(1):21-7. doi: 10.1016/s0168-9525(99)01857-0. Trends Genet. 2000. PMID: 10637627 Review.
-
The Caenorhabditis elegans cuticle and precuticle: a model for studying dynamic apical extracellular matrices in vivo.Genetics. 2024 Aug 7;227(4):iyae072. doi: 10.1093/genetics/iyae072. Genetics. 2024. PMID: 38995735 Free PMC article. Review.
Cited by
-
Epidermal PAR-6 and PKC-3 are essential for larval development of C. elegans and organize non-centrosomal microtubules.Elife. 2020 Dec 10;9:e62067. doi: 10.7554/eLife.62067. Elife. 2020. PMID: 33300872 Free PMC article.
-
Defects in the C. elegans acyl-CoA synthase, acs-3, and nuclear hormone receptor, nhr-25, cause sensitivity to distinct, but overlapping stresses.PLoS One. 2014 Mar 20;9(3):e92552. doi: 10.1371/journal.pone.0092552. eCollection 2014. PLoS One. 2014. PMID: 24651852 Free PMC article.
-
High-content phenotypic analysis of a C. elegans recombinant inbred population identifies genetic and molecular regulators of lifespan.Cell Rep. 2024 Oct 22;43(10):114836. doi: 10.1016/j.celrep.2024.114836. Epub 2024 Oct 4. Cell Rep. 2024. PMID: 39368088 Free PMC article.
-
Culture and Assay of Large-Scale Mixed-Stage Caenorhabditis elegans Populations.J Vis Exp. 2021 May 5;(171):10.3791/61453. doi: 10.3791/61453. J Vis Exp. 2021. PMID: 34028439 Free PMC article.
-
Analysis of a lin-42/period Null Allele Implicates All Three Isoforms in Regulation of Caenorhabditis elegans Molting and Developmental Timing.G3 (Bethesda). 2016 Dec 7;6(12):4077-4086. doi: 10.1534/g3.116.034165. G3 (Bethesda). 2016. PMID: 27729432 Free PMC article.
References
-
- Aguinaldo A. M., Turbeville J. M., Linford L. S., Rivera M. C., Garey J. R., Raff R. A., Lake J. A. Evidence for a clade of nematodes, arthropods and other moulting animals. Nature. 1997;387:489–493. - PubMed
-
- Altschul S. F., Gish W., Miller W., Myers E. W., Lipman D. J. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. - PubMed
-
- Asahina M., Ishihara T., Jindra M., Kohara Y., Katsura I., Hirose S. The conserved nuclear receptor Ftz-F1 is required for embryogenesis, moulting and reproduction in Caenorhabditis elegans. Genes Cells. 2000;5:711–723. - PubMed
-
- Bessereau J. L., Wright A., Williams D. C., Schuske K., Davis M. W., Jorgensen E. M. Mobilization of a Drosophila transposon in the Caenorhabditis elegans germ line. Nature. 2001;413:70–74. - PubMed
-
- Bhattacharjee A., Bansal M. Collagen structure: the Madras triple helix and the current scenario. IUBMB Life. 2005;57:161–172. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
