

Contrasting models for kinetochore microtubule attachment in mammalian cells
- PMID: 20336345
- PMCID: PMC2883615
- DOI: 10.1007/s00018-010-0322-x
Contrasting models for kinetochore microtubule attachment in mammalian cells
Abstract
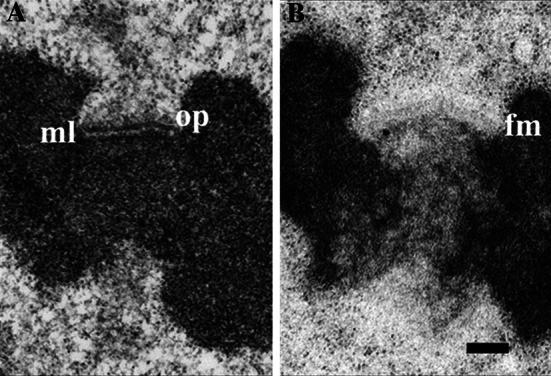
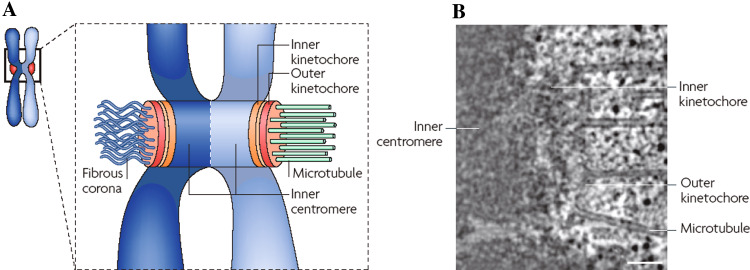
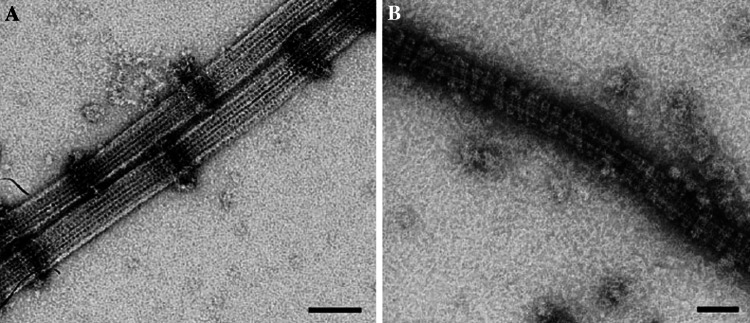
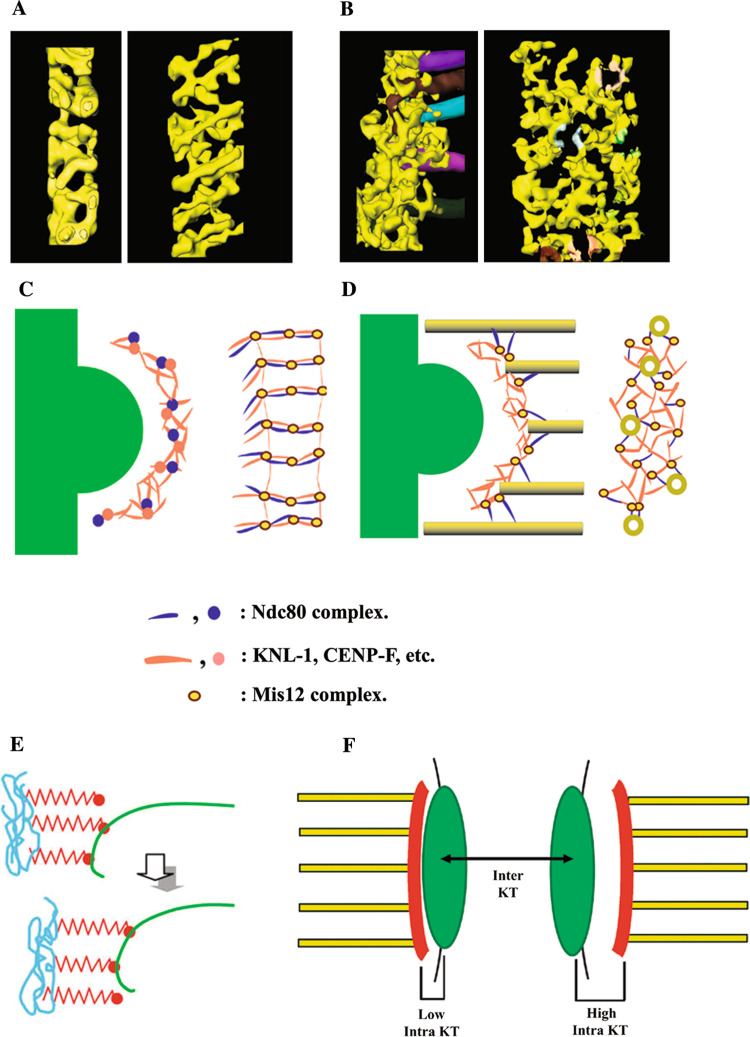
Kinetochore function is mediated through its interaction with microtubule plus ends embedded in the kinetochore outer plate. Here, we compare and evaluate current models for kinetochore microtubule attachment, beginning with a brief review of the molecular, biochemical, cellular, and structural studies upon which these models are based. The majority of these studies strongly support a model in which the kinetochore outer plate is a network of fibers that form multiple weak attachments to each microtubule, chiefly through the Ndc80 complex. Multiple weak attachments enable kinetochores to remain attached to microtubule plus ends that are continually growing and shrinking. It is unlikely that rings or "kinetochore fibrils" have a significant role in kinetochore microtubule attachment, but such entities could have a role in stabilizing attachment, modifying microtubule dynamics, and harnessing the energy released from microtubule disassembly. It is currently unclear whether kinetochores control and coordinate the dynamics of individual kinetochore microtubules.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
