

Arabidopsis thaliana XRN2 is required for primary cleavage in the pre-ribosomal RNA
- PMID: 20338880
- PMCID: PMC2910052
- DOI: 10.1093/nar/gkq172
Arabidopsis thaliana XRN2 is required for primary cleavage in the pre-ribosomal RNA
Abstract
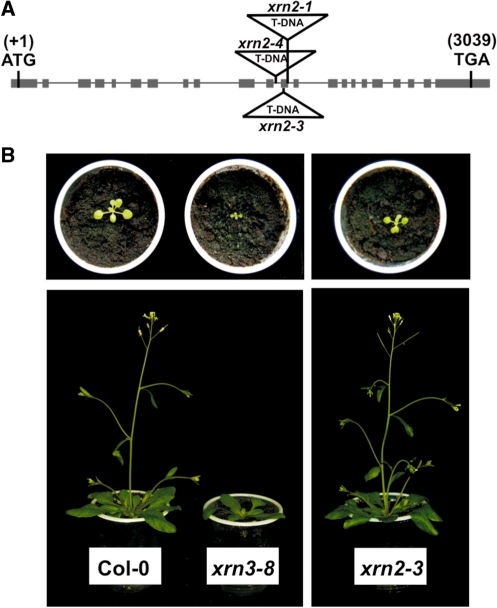
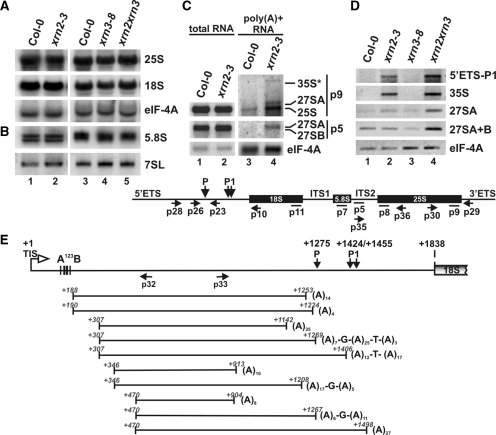
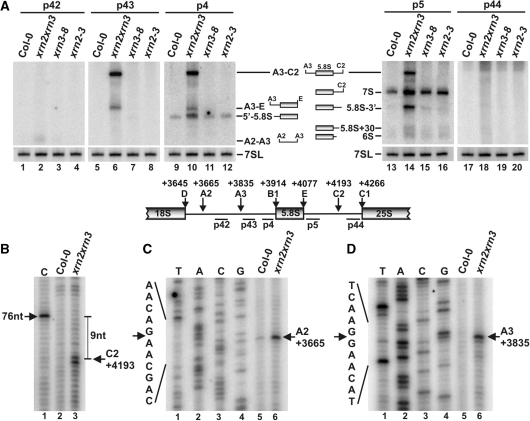
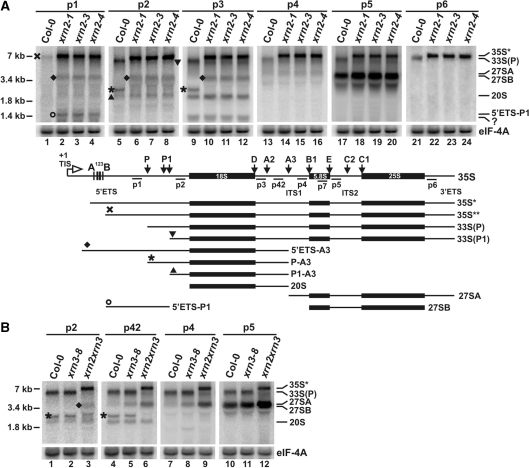
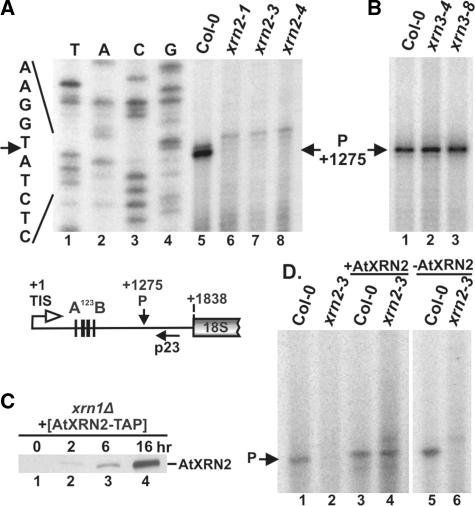
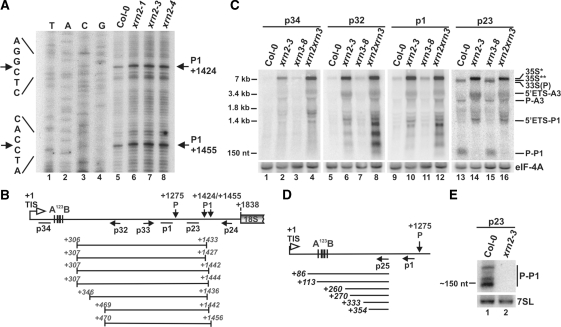
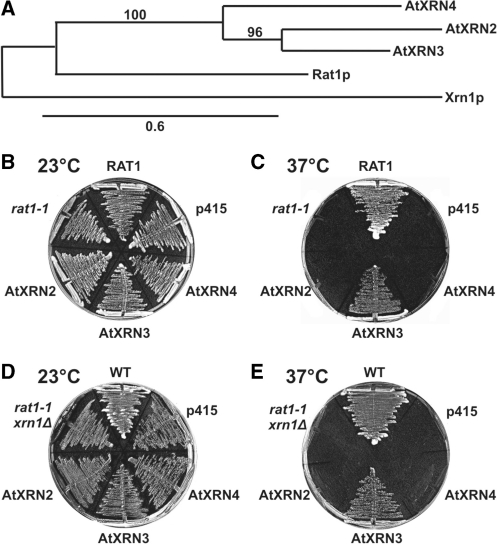
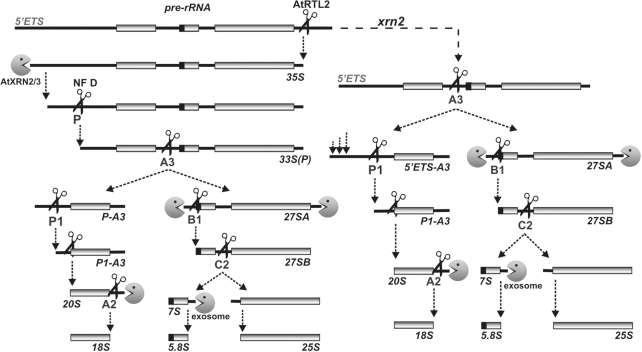
Three Rat1/Xrn2 homologues exist in Arabidopsis thaliana: nuclear AtXRN2 and AtXRN3, and cytoplasmic AtXRN4. The latter has a role in degrading 3' products of miRNA-mediated mRNA cleavage, whereas all three proteins act as endogenous post-transcriptional gene silencing suppressors. Here we show that, similar to yeast nuclear Rat1, AtXRN2 has a role in ribosomal RNA processing. The lack of AtXRN2, however, does not result in defective formation of rRNA 5'-ends but inhibits endonucleolytic cleavage at the primary site P in the pre-rRNA resulting in the accumulation of the 35S* precursor. This does not lead to a decrease in mature rRNAs, as additional cleavages occur downstream of site P. Supplementing a P-site cleavage-deficient xrn2 plant extract with the recombinant protein restores processing activity, indicating direct participation of AtXRN2 in this process. Our data suggest that the 5' external transcribed spacer is shortened by AtXRN2 prior to cleavage at site P and that this initial exonucleolytic trimming is required to expose site P for subsequent endonucleolytic processing by the U3 snoRNP complex. We also show that some rRNA precursors and excised spacer fragments that accumulate in the absence of AtXRN2 and AtXRN3 are polyadenylated, indicating that these nucleases contribute to polyadenylation-dependent nuclear RNA surveillance.
Figures
References
-
- Garneau NL, Wilusz J, Wilusz CJ. The highways and byways of mRNA decay. Nat. Rev. Mol. Cell. Biol. 2007;8:113–126. - PubMed
-
- Houseley J, Tollervey D. The many pathways of RNA degradation. Cell. 2009;136:763–776. - PubMed
-
- Souret FF, Kastenmayer JP, Green PJ. AtXRN4 degrades mRNA in Arabidopsis and its substrates include selected miRNA targets. Mol. Cell. 2004;15:173–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
