

Recent advances in understanding the assembly and repair of photosystem II
- PMID: 20338950
- PMCID: PMC2889791
- DOI: 10.1093/aob/mcq059
Recent advances in understanding the assembly and repair of photosystem II
Abstract
Background: Photosystem II (PSII) is the light-driven water:plastoquinone oxidoreductase of oxygenic photosynthesis and is found in the thylakoid membrane of chloroplasts and cyanobacteria. Considerable attention is focused on how PSII is assembled in vivo and how it is repaired following irreversible damage by visible light (so-called photoinhibition). Understanding these processes might lead to the development of plants with improved growth characteristics especially under conditions of abiotic stress.
Scope: Here we summarize recent results on the assembly and repair of PSII in cyanobacteria, which are excellent model organisms to study higher plant photosynthesis.
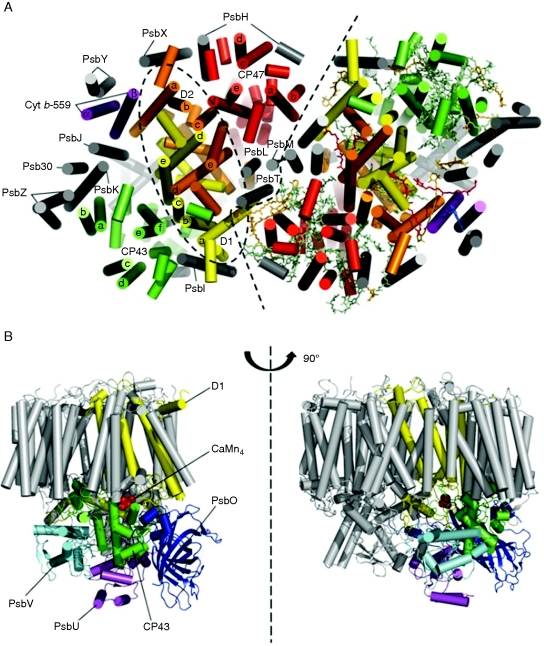
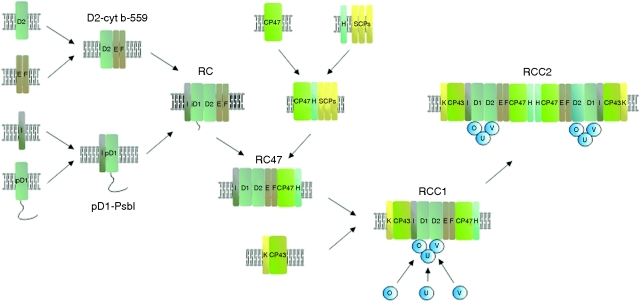
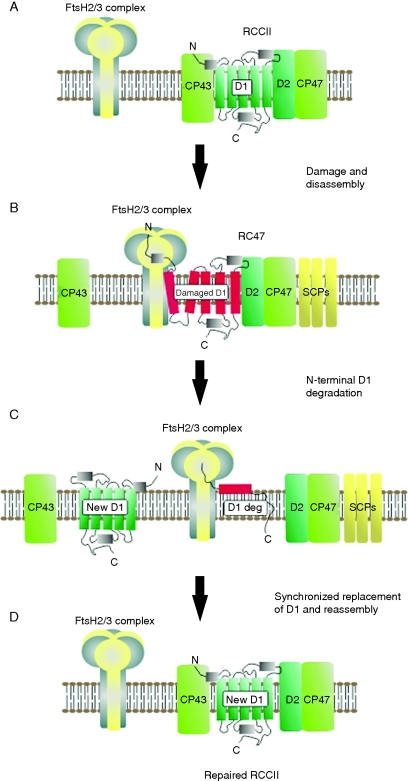
Conclusions: Assembly of PSII is highly co-ordinated and proceeds through a number of distinct assembly intermediates. Associated with these assembly complexes are proteins that are not found in the final functional PSII complex. Structural information and possible functions are beginning to emerge for several of these 'assembly' factors, notably Ycf48/Hcf136, Psb27 and Psb28. A number of other auxiliary proteins have been identified that appear to have evolved since the divergence of chloroplasts and cyanobacteria. The repair of PSII involves partial disassembly of the damaged complex, the selective replacement of the damaged sub-unit (predominantly the D1 sub-unit) by a newly synthesized copy, and reassembly. It is likely that chlorophyll released during the repair process is temporarily stored by small CAB-like proteins (SCPs). A model is proposed in which damaged D1 is removed in Synechocystis sp. PCC 6803 by a hetero-oligomeric complex composed of two different types of FtsH sub-unit (FtsH2 and FtsH3), with degradation proceeding from the N-terminus of D1 in a highly processive reaction. It is postulated that a similar mechanism of D1 degradation also operates in chloroplasts. Deg proteases are not required for D1 degradation in Synechocystis 6803 but members of this protease family might play a supplementary role in D1 degradation in chloroplasts under extreme conditions.
Figures
References
-
- Adam Z, Clarke AK. Cutting edge of chloroplast proteolysis. Trends in Plant Science. 2002;7:451–456. - PubMed
-
- Adam Z, Zaltsman A, Sinvany-Villalobo G, Sakamoto W. FtsH proteases in chloroplasts and cyanobacteria. Physiologia Plantarum. 2005;123:386–390.
-
- Adir N, Zer H, Shochat S, Ohad I. Photoinhibition – a historical perspective. Photosynthesis Research. 2003;76:343–370. - PubMed
-
- Akiyama Y, Ito K. Reconstitution of membrane proteolysis by FtsH. Journal of Biological Chemistry. 2003;278:18146–18153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
