

3D-Reconstructions and Virtual 4D-Visualization to Study Metamorphic Brain Development in the Sphinx Moth Manduca Sexta
- PMID: 20339481
- PMCID: PMC2845058
- DOI: 10.3389/fnsys.2010.00007
3D-Reconstructions and Virtual 4D-Visualization to Study Metamorphic Brain Development in the Sphinx Moth Manduca Sexta
Abstract
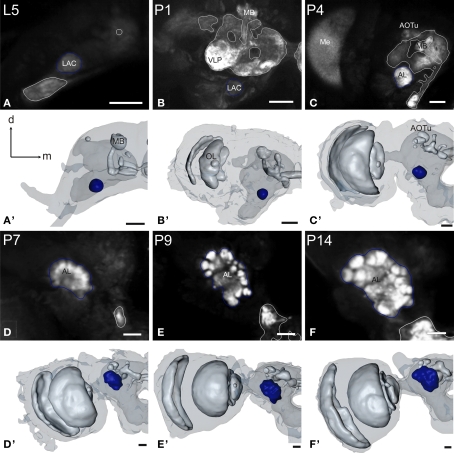
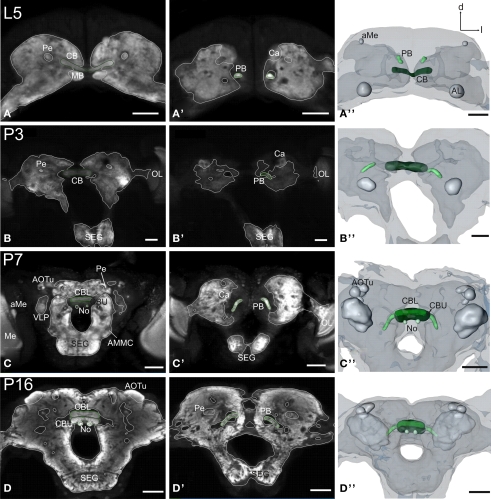
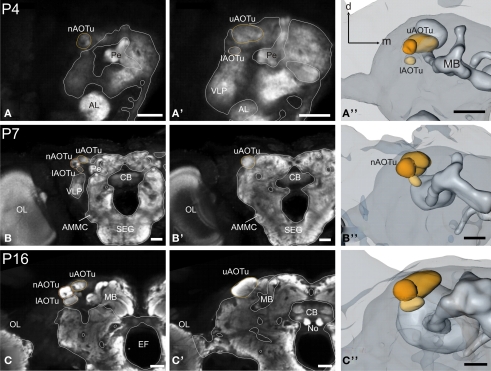
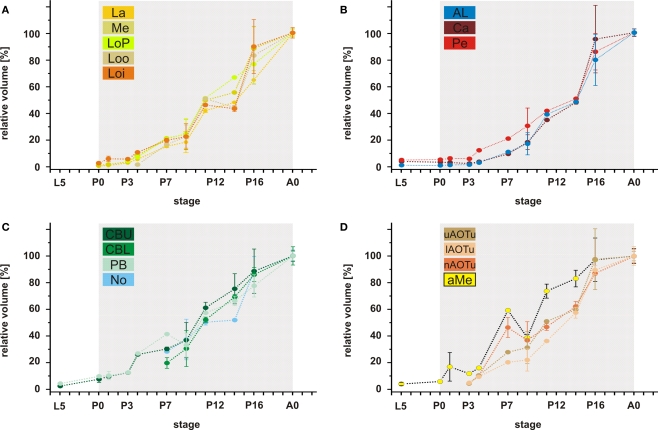
DURING METAMORPHOSIS, THE TRANSITION FROM THE LARVA TO THE ADULT, THE INSECT BRAIN UNDERGOES CONSIDERABLE REMODELING: new neurons are integrated while larval neurons are remodeled or eliminated. One well acknowledged model to study metamorphic brain development is the sphinx moth Manduca sexta. To further understand mechanisms involved in the metamorphic transition of the brain we generated a 3D standard brain based on selected brain areas of adult females and 3D reconstructed the same areas during defined stages of pupal development. Selected brain areas include for example mushroom bodies, central complex, antennal- and optic lobes. With this approach we eventually want to quantify developmental changes in neuropilar architecture, but also quantify changes in the neuronal complement and monitor the development of selected neuronal populations. Furthermore, we used a modeling software (Cinema 4D) to create a virtual 4D brain, morphing through its developmental stages. Thus the didactical advantages of 3D visualization are expanded to better comprehend complex processes of neuropil formation and remodeling during development. To obtain datasets of the M. sexta brain areas, we stained whole brains with an antiserum against the synaptic vesicle protein synapsin. Such labeled brains were then scanned with a confocal laser scanning microscope and selected neuropils were reconstructed with the 3D software AMIRA 4.1.
Keywords: Manduca; animation; brain; development; digital neuroanatomy; insect; neuropil.
Figures




References
-
- Amos T. M., Gelman D. B., Mesce K. A. (1996). Steroid hormone fluctuations regulate ganglionic fusion during metamorphosis of the moth Manduca sexta. J. Insect Physiol. 42, 579–591 10.1016/0022-1910(95)00127-1 - DOI
-
- Amos T. M., Mesce K. A. (1994). Reorganization of the ventral nerve cord in the moth Manduca sexta (L.) (Lepidoptera:Sphingidae). Int. J. Insect Morphol. Embryol. 23, 21–37 10.1016/0020-7322(94)90013-2 - DOI
-
- Bell R. A., Joachim F. A. (1978). Techniques for rearing laboratory colonies of the tobacco hornworm, Manduca sexta, and pink ballworms. Ann. Entomol. Soc. Am. 69, 365–373
-
- Bullock T. H., Horridge G. A. (1965). Structure and Function in the Nervous Systems of Invertebrates, Vol. 2 San Francisco, Freeman
LinkOut - more resources
Full Text Sources