

The elegans of spindle assembly
- PMID: 20339898
- PMCID: PMC2883083
- DOI: 10.1007/s00018-010-0324-8
The elegans of spindle assembly
Abstract
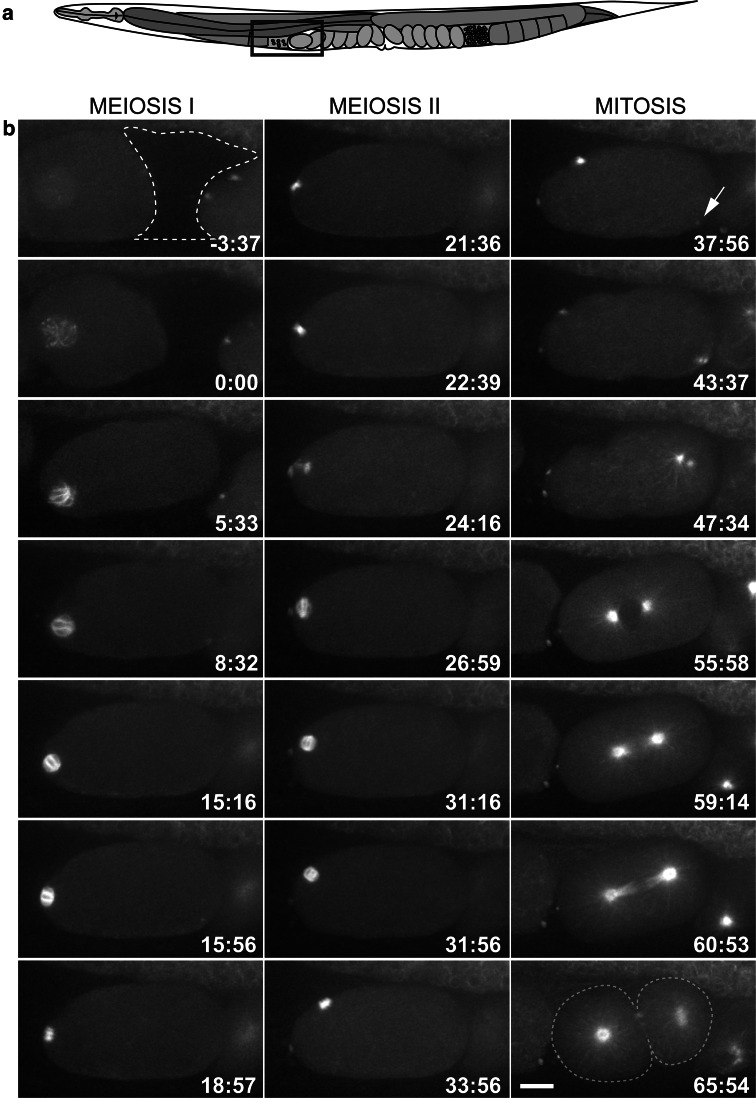
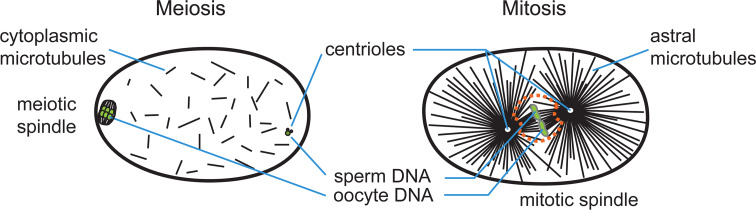
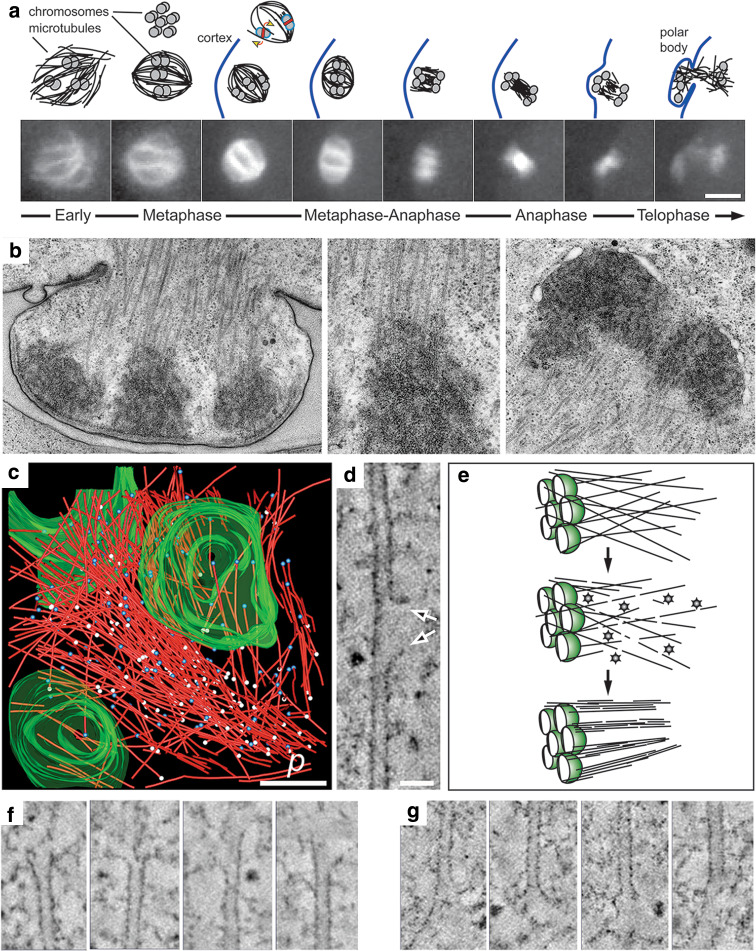
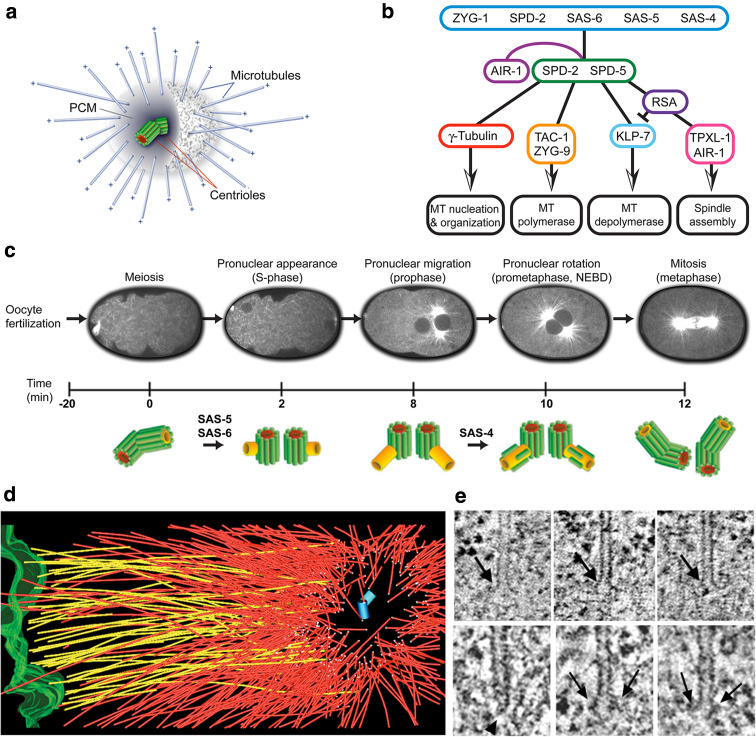
The Caenorhabditis elegans one-cell embryo is a powerful system in which to study microtubule organization because this large cell assembles both meiotic and mitotic spindles within the same cytoplasm over the course of 1 h in a stereotypical manner. The fertilized oocyte assembles two consecutive acentrosomal meiotic spindles that function to reduce the replicated maternal diploid set of chromosomes to a single-copy haploid set. The resulting maternal DNA then unites with the paternal DNA to form a zygotic diploid complement, around which a centrosome-based mitotic spindle forms. The early C. elegans embryo is amenable to live-cell imaging and electron tomography, permitting a detailed structural comparison of the meiotic and mitotic modes of spindle assembly.
Figures
Similar articles
-
MEI-1/katanin is required for translocation of the meiosis I spindle to the oocyte cortex in C elegans.Dev Biol. 2003 Aug 1;260(1):245-59. doi: 10.1016/s0012-1606(03)00216-1. Dev Biol. 2003. PMID: 12885567
-
Katanin disrupts the microtubule lattice and increases polymer number in C. elegans meiosis.Curr Biol. 2006 Oct 10;16(19):1944-9. doi: 10.1016/j.cub.2006.08.029. Curr Biol. 2006. PMID: 17027492
-
Katanin controls mitotic and meiotic spindle length.J Cell Biol. 2006 Dec 18;175(6):881-91. doi: 10.1083/jcb.200608117. J Cell Biol. 2006. PMID: 17178907 Free PMC article.
-
Oocyte Meiotic Spindle Assembly and Function.Curr Top Dev Biol. 2016;116:65-98. doi: 10.1016/bs.ctdb.2015.11.031. Epub 2016 Jan 23. Curr Top Dev Biol. 2016. PMID: 26970614 Free PMC article. Review.
-
Centrosomes and the art of mitotic spindle maintenance.Int Rev Cell Mol Biol. 2014;313:179-217. doi: 10.1016/B978-0-12-800177-6.00006-2. Int Rev Cell Mol Biol. 2014. PMID: 25376493 Review.
Cited by
-
It takes two (centrioles) to tango.Reproduction. 2019 Feb;157(2):R33-R51. doi: 10.1530/REP-18-0350. Reproduction. 2019. PMID: 30496124 Free PMC article. Review.
-
A semi-dominant mutation in the general splicing factor SF3a66 causes anterior-posterior axis reversal in one-cell stage C. elegans embryos.PLoS One. 2014 Sep 4;9(9):e106484. doi: 10.1371/journal.pone.0106484. eCollection 2014. PLoS One. 2014. PMID: 25188372 Free PMC article.
-
KLP-7 acts through the Ndc80 complex to limit pole number in C. elegans oocyte meiotic spindle assembly.J Cell Biol. 2015 Sep 14;210(6):917-32. doi: 10.1083/jcb.201412010. J Cell Biol. 2015. PMID: 26370499 Free PMC article.
-
The non-catalytic domains of Drosophila katanin regulate its abundance and microtubule-disassembly activity.PLoS One. 2015 Apr 17;10(4):e0123912. doi: 10.1371/journal.pone.0123912. eCollection 2015. PLoS One. 2015. PMID: 25886649 Free PMC article.
-
Meiotic kinetochores get pushed aside by a CLS act.Nat Cell Biol. 2010 Sep;12(9):849-51. doi: 10.1038/ncb0910-849. Epub 2010 Aug 22. Nat Cell Biol. 2010. PMID: 20729839
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
