

doi: 10.1111/j.1742-4658.2010.07623.x.
Epub 2010 Mar 22.
Effects of a novel arginine methyltransferase inhibitor on T-helper cell cytokine production
Affiliations
- PMID: 20345902
- PMCID: PMC2903848
- DOI: 10.1111/j.1742-4658.2010.07623.x
Item in Clipboard
Effects of a novel arginine methyltransferase inhibitor on T-helper cell cytokine production
FEBS J.
2010 May.
Abstract
The protein arginine methyltransferase (PRMT) family of enzymes catalyzes the transfer of methyl groups from S-adenosylmethionine to the guanidino nitrogen atom of peptidylarginine to form monomethylarginine or dimethylarginine. We created several less polar analogs of the specific PRMT inhibitor arginine methylation inhibitor-1, and one such compound was found to have improved PRMT inhibitory activity over the parent molecule. The newly identified PRMT inhibitor modulated T-helper-cell function and thus may serve as a lead for further inhibitors useful for the treatment of immune-mediated disease.
Figures
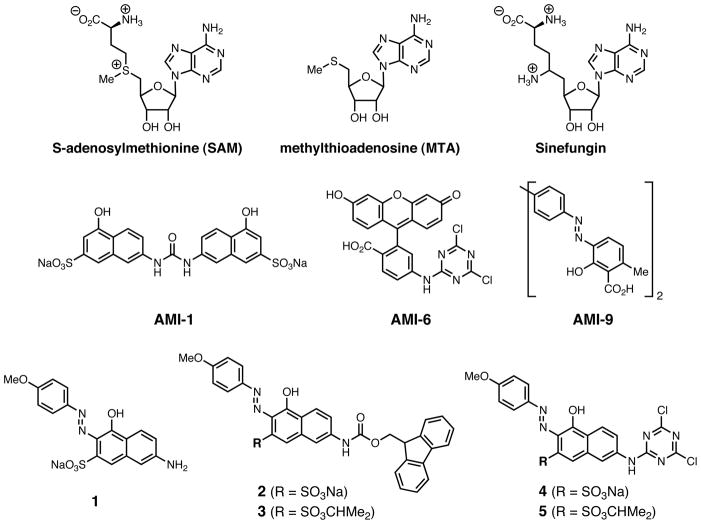
Chemical structures of SAM, MTA, Sinefungin, AMI-1, AMI-6, AMI-9, and compounds 1–5.
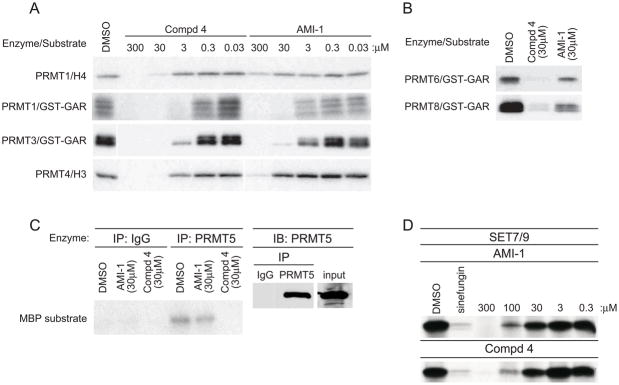
a) in vitro methylation reactions with recombinant GST-PRMT1, GST-PRMT3, and GST-PRMT4 with indicated substrate and [3H]SAM in the presence of increasing concentrations of AMI-1 or 4. b) in vitro methylation reactions with recombinant GST-PRMT6 or GST-PRMT8 together with GST-GAR and [3H]SAM in the presence of 30μM AMI-1 or 4. c) Immunoprecipitated PRMT5 or isotype control from 293 cell extracts was subjected to in vitro methylation reactions using the indicated concentrations of AMI-1 or 4 and MBP as substrate (left panel). Reaction inputs were determined by immunoblotting with PRMT5 antisera (right panel). d) in vitro methylation reactions with recombinant Set7/9 with calf thymus histones as substrate and [3H]SAM in the presence of increasing concentrations of AMI-1 or 4. Data are representative of at least three independent experiments.
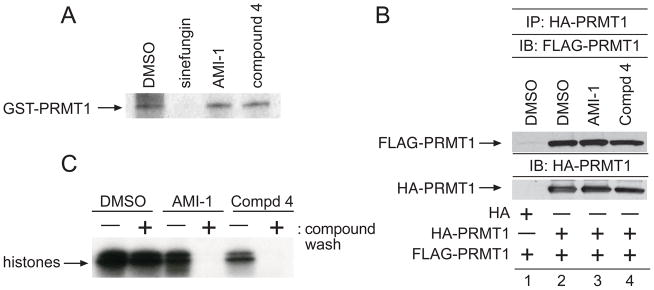
a) GST-PRMT1 was UV-crosslinked to [3H]SAM in the presence of DMSO, sinefugin (100μM), AMI-1 (100μM), or 4 (100μM), separated by SDS-PAGE, and visualized by fluorography b) 293T cells were transfected with HA-PRMT1 or FLAG-PRMT1. Lysates from the FLAG-PRMT1 transfection were incubated with HA-PRMT1 immunoprecipitates in the presence of DMSO (lane 2), AMI-1 (100μM, lane 3), or 4 (100μM, lane 4), resolved by SDS-PAGE, and the immunoblot was incubated with an antibody to FLAG. Reprobing the immunoblot with an antibody to HA demonstrated equal loading. Specificity of the HA-PRMT1/FLAG-PRMT interaction was determined by incubating immunoprecipitates from vector only transfected cells with FLAG-NIP45 lysates c) Incubations of GST-PRMT1 glutathione beads with DMSO, AMI-1, or 4 were divided into two aliquots. Bead aliquots were washed either in the presence (+) or absence (−) of indicated compounds. Washed aliquots were immediately subjected to in vitro methylation assays using calf thymus histones. Data are representative of three independent experiments.
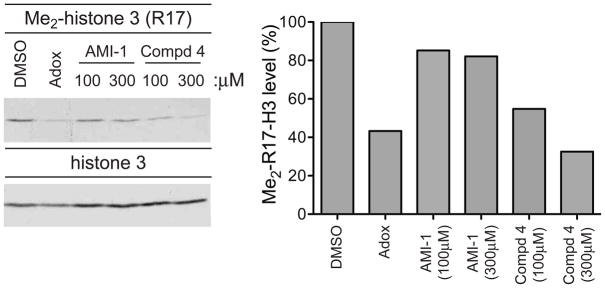
293T cells were treated with DMSO, AMI-1 (100μM or 300μM), 4 (100μM or 300μM), or Adox (20μM) for 24 hrs. Histone extracts were immunoblotted for H3R17 methylation (left panel). Quantification of the methylation levels of compound-treated samples relative to vehicle-treated samples is depicted in the right panel. Data are representative of three independent experiments.
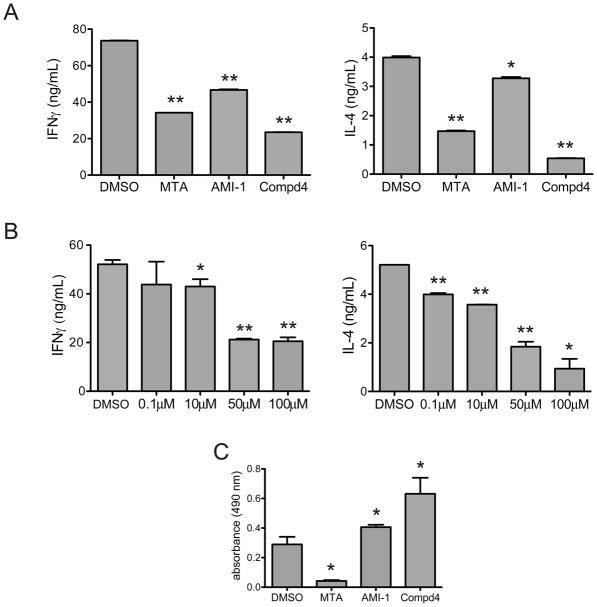
a) Th1 or Th2 were stimulated with plate-bound anti-CD3 in the presence of DMSO, MTA (100μM), AMI-1 (100μM), and 4 (100μM). Supernatants were analyzed by ELISA to determine Th1 production of IFNγ (left panel) or Th2 production of IL-4 (right panel). b) Th1 or Th2 cells were stimulated with plate-bound anti-CD3 in the presence of DMSO or varying concentrations of 4, and IFNγ (left panel, Th1 cells) or IL-4 (right panel, Th2 cells) levels were determined by ELISA. c) T helper cells were stimulated with plate-bound anti-CD3 in the presence of DMSO, MTA, AMI-1, or 4. Cellular proliferation was determined by the MTS assay. * p < 0.05, ** p < 0.01. Data are representative of at least three independent experiments.
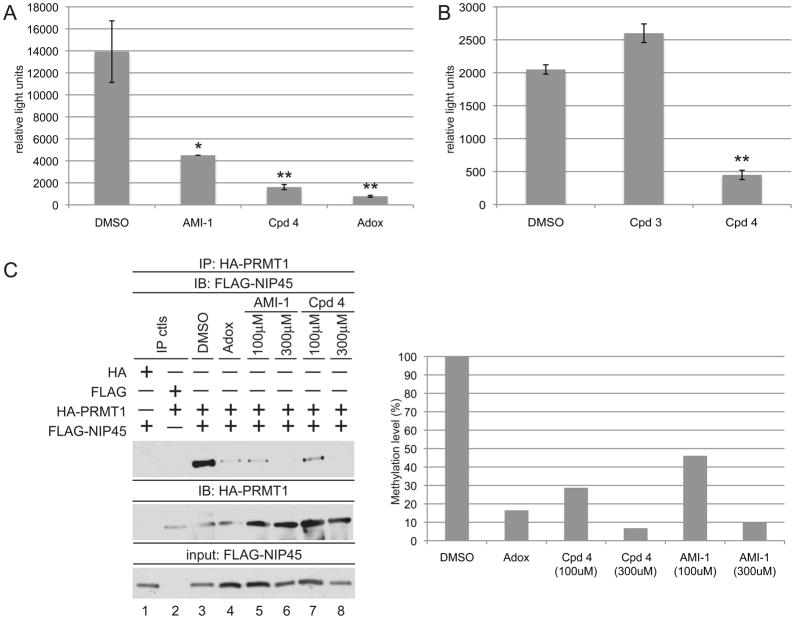
a) Jurkat cells were transfected with the IL-4 luciferase reporter (3μg) along with TK-Renilla luciferase vector (10ng) as an internal control. Transfectants were pretreated with DMSO, AMI-1 (100μM), 4 (100μM), or Adox (20μM) for 18hrs prior to 6hr stimulation with PMA/Ionomycin. Luciferase values were calculated relative to TK-Renilla luciferase internal controls. Similar results were obtained in at least three independent experiments. * p < 0.05, ** p < 0.01. b) same as in a) except cells were treated with 3 (100μM) or 4 (100μM). ** p < 0.01 c) 293T cells transfected with HA-PRMT1 and FLAG-NIP45 expression vectors were treated with DMSO (lane3), Adox (20μM, lane 4), AMI-1 (100μM, lanes 5–6), or 4 (100μM, lanes 7–8). Lysates were immunoprecipitated with anti-HA agarose, and immunoprecipitates were probed for FLAG-NIP45 using an antibody recognizing the FLAG epitope (top panel). HA-PRMT immunoprecipitate levels were evaluated by reblotting with an antibody to HA (middle panel). Bottom panel demonstrates FLAG-NIP45 input levels. Quantification of FLAG-NIP45/HA-PRMT association levels are depicted relative to the DMSO treated sample (right panel). Data are representative of three independent experiments.
References
-
- Lin WJ, Gary JD, Yang MC, Clarke S, Herschman HR. The mammalian immediate-early TIS21 protein and the leukemia-associated BTG1 protein interact with a protein-arginine N-methyltransferase. J Biol Chem. 1996;271:15034–15044. - PubMed
-
- Bedford MT, Richard S. Arginine methylation an emerging regulator of protein function. Mol Cell. 2005;18:263–272. - PubMed
-
- Krause CD, Yang ZH, Kim YS, Lee JH, Cook JR, Pestka S. Protein arginine methyltransferases: Evolution and assessment of their pharmacological and therapeutic potential. Pharmacol Ther. 2007;113:50–87. - PubMed
-
- Lee DY, Ianculescu I, Purcell D, Zhang X, Cheng X, Stallcup MR. Surface-scanning mutational analysis of protein arginine methyltransferase 1: roles of specific amino acids in methyltransferase substrate specificity, oligomerization, and coactivator function. Mol Endocrinol. 2007;21:1381–1393. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
