

Social cognition and the evolution of language: constructing cognitive phylogenies
- PMID: 20346756
- PMCID: PMC4415479
- DOI: 10.1016/j.neuron.2010.03.011
Social cognition and the evolution of language: constructing cognitive phylogenies
Abstract
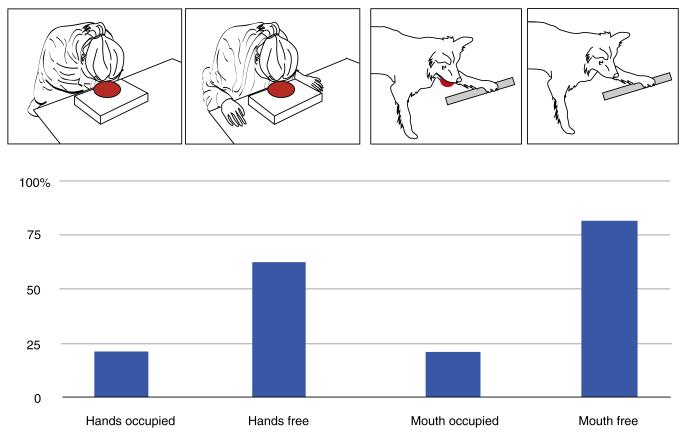
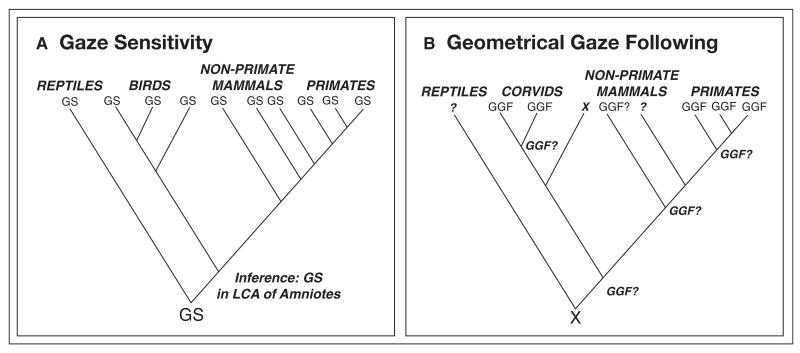
Human language and social cognition are closely linked: advanced social cognition is necessary for children to acquire language, and language allows forms of social understanding (and, more broadly, culture) that would otherwise be impossible. Both "language" and "social cognition" are complex constructs, involving many independent cognitive mechanisms, and the comparative approach provides a powerful route to understanding the evolution of such mechanisms. We provide a broad comparative review of mechanisms underlying social intelligence in vertebrates, with the goal of determining which human mechanisms are broadly shared, which have evolved in parallel in other clades, and which, potentially, are uniquely developed in our species. We emphasize the importance of convergent evolution for testing hypotheses about neural mechanisms and their evolution.
(c) 2010 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
Primate social cognition: uniquely primate, uniquely social, or just unique?Neuron. 2010 Mar 25;65(6):815-30. doi: 10.1016/j.neuron.2010.03.010. Neuron. 2010. PMID: 20346757 Review.
-
Embodied cognitive evolution and the cerebellum.Philos Trans R Soc Lond B Biol Sci. 2012 Aug 5;367(1599):2097-107. doi: 10.1098/rstb.2012.0112. Philos Trans R Soc Lond B Biol Sci. 2012. PMID: 22734053 Free PMC article.
-
Genes, language, cognition, and culture: towards productive inquiry.Hum Biol. 2011 Apr;83(2):323-9. doi: 10.3378/027.083.0210. Hum Biol. 2011. PMID: 21615292
-
Humans have evolved specialized skills of social cognition: the cultural intelligence hypothesis.Science. 2007 Sep 7;317(5843):1360-6. doi: 10.1126/science.1146282. Science. 2007. PMID: 17823346
-
The evolution of the capacity for language: the ecological context and adaptive value of a process of cognitive hijacking.Philos Trans R Soc Lond B Biol Sci. 2018 Apr 5;373(1743):20170052. doi: 10.1098/rstb.2017.0052. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 29440518 Free PMC article. Review.
Cited by
-
Of mice, birds, and men: the mouse ultrasonic song system has some features similar to humans and song-learning birds.PLoS One. 2012;7(10):e46610. doi: 10.1371/journal.pone.0046610. Epub 2012 Oct 10. PLoS One. 2012. PMID: 23071596 Free PMC article.
-
Commentary on: Labels, cognomes, and cyclic computation: an ethological perspective.Front Psychol. 2015 Jun 11;6:784. doi: 10.3389/fpsyg.2015.00784. eCollection 2015. Front Psychol. 2015. PMID: 26113831 Free PMC article. No abstract available.
-
Yes, you can? A speaker's potency to act upon his words orchestrates early neural responses to message-level meaning.PLoS One. 2013 Jul 24;8(7):e69173. doi: 10.1371/journal.pone.0069173. Print 2013. PLoS One. 2013. PMID: 23894425 Free PMC article. Clinical Trial.
-
Universal Grammar and Biological Variation: An EvoDevo Agenda for Comparative Biolinguistics.Biol Theory. 2014;9(2):122-134. doi: 10.1007/s13752-014-0164-0. Epub 2014 Mar 15. Biol Theory. 2014. PMID: 24955079 Free PMC article.
-
Ethics trumps resources in women's and men's evaluations of potential mates and competitors.Sci Rep. 2025 Jul 2;15(1):22597. doi: 10.1038/s41598-025-06611-8. Sci Rep. 2025. PMID: 40596369 Free PMC article.
References
-
- Agnetta B, Hare B, Tomasello M. Cues to food location that domestic dogs (Canis familiaris) of different ages do and do not use. Anim. Cogn. 2000;3:107–112.
-
- Aitken PG. Cortical control of conditioned and spontaneous vocal behavior in rhesus monkeys. Brain Lang. 1981;13:171–184. - PubMed
-
- Anderson JR, Montant M, Schmitt D. Rhesus monkeys fail to use gaze direction as an experimenter-given cue in an object-choice task. Behav. Processes. 1996;37:47–55. - PubMed
-
- Arbib MA. From monkey-like action recognition to human language: an evolutionary framework for neurolinguistics. Behav. Brain Sci. 2005;28:105–124. discussion 125–167. - PubMed
-
- Baptista LF, Petrinovich L. Song development in the white-crowned sparrow: social factors and sex differences. Anim. Behav. 1986;34:1359–1371.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
