

Postnatal deamidation of 4E-BP2 in brain enhances its association with raptor and alters kinetics of excitatory synaptic transmission
- PMID: 20347422
- PMCID: PMC2861547
- DOI: 10.1016/j.molcel.2010.02.022
Postnatal deamidation of 4E-BP2 in brain enhances its association with raptor and alters kinetics of excitatory synaptic transmission
Abstract
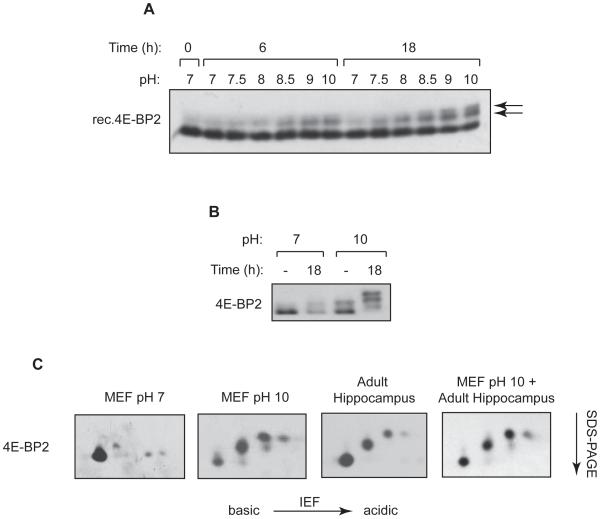
The eIF4E-binding proteins (4E-BPs) repress translation initiation by preventing eIF4F complex formation. Of the three mammalian 4E-BPs, only 4E-BP2 is enriched in the mammalian brain and plays an important role in synaptic plasticity and learning and memory formation. Here we describe asparagine deamidation as a brain-specific posttranslational modification of 4E-BP2. Deamidation is the spontaneous conversion of asparagines to aspartates. Two deamidation sites were mapped to an asparagine-rich sequence unique to 4E-BP2. Deamidated 4E-BP2 exhibits increased binding to the mammalian target of rapamycin (mTOR)-binding protein raptor, which effects its reduced association with eIF4E. 4E-BP2 deamidation occurs during postnatal development, concomitant with the attenuation of the activity of the PI3K-Akt-mTOR signaling pathway. Expression of deamidated 4E-BP2 in 4E-BP2(-/-) neurons yielded mEPSCs exhibiting increased charge transfer with slower rise and decay kinetics relative to the wild-type form. 4E-BP2 deamidation may represent a compensatory mechanism for the developmental reduction of PI3K-Akt-mTOR signaling.
(c) 2010 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
The Unified Theory of Neurodegeneration Pathogenesis Based on Axon Deamidation.Int J Mol Sci. 2025 Apr 27;26(9):4143. doi: 10.3390/ijms26094143. Int J Mol Sci. 2025. PMID: 40362380 Free PMC article. Review.
-
Repair of isoaspartate formation modulates the interaction of deamidated 4E-BP2 with mTORC1 in brain.J Biol Chem. 2010 Jun 18;285(25):19402-8. doi: 10.1074/jbc.M110.120774. Epub 2010 Apr 27. J Biol Chem. 2010. PMID: 20424163 Free PMC article.
-
Raptor-Mediated Proteasomal Degradation of Deamidated 4E-BP2 Regulates Postnatal Neuronal Translation and NF-κB Activity.Cell Rep. 2019 Dec 10;29(11):3620-3635.e7. doi: 10.1016/j.celrep.2019.11.023. Cell Rep. 2019. PMID: 31825840 Free PMC article.
-
Selective regulation of GluA subunit synthesis and AMPA receptor-mediated synaptic function and plasticity by the translation repressor 4E-BP2 in hippocampal pyramidal cells.J Neurosci. 2013 Jan 30;33(5):1872-86. doi: 10.1523/JNEUROSCI.3264-12.2013. J Neurosci. 2013. PMID: 23365227 Free PMC article.
-
Signalling to eIF4E in cancer.Biochem Soc Trans. 2015 Oct;43(5):763-72. doi: 10.1042/BST20150126. Biochem Soc Trans. 2015. PMID: 26517881 Free PMC article. Review.
Cited by
-
Viral pseudo-enzymes activate RIG-I via deamidation to evade cytokine production.Mol Cell. 2015 Apr 2;58(1):134-46. doi: 10.1016/j.molcel.2015.01.036. Epub 2015 Mar 5. Mol Cell. 2015. PMID: 25752576 Free PMC article.
-
Transcriptional induction of 4E-BP3 prolongs translation repression.Cell Cycle. 2016 Dec 16;15(24):3325-3326. doi: 10.1080/15384101.2016.1224786. Epub 2016 Aug 26. Cell Cycle. 2016. PMID: 27565030 Free PMC article. No abstract available.
-
Insulin signaling controls neurotransmission via the 4eBP-dependent modification of the exocytotic machinery.Elife. 2016 Aug 15;5:e16807. doi: 10.7554/eLife.16807. Elife. 2016. PMID: 27525480 Free PMC article.
-
Extensive genetic differentiation between recently evolved sympatric Arctic charr morphs.Ecol Evol. 2019 Sep 12;9(19):10964-10983. doi: 10.1002/ece3.5516. eCollection 2019 Oct. Ecol Evol. 2019. PMID: 31641448 Free PMC article.
-
The Unified Theory of Neurodegeneration Pathogenesis Based on Axon Deamidation.Int J Mol Sci. 2025 Apr 27;26(9):4143. doi: 10.3390/ijms26094143. Int J Mol Sci. 2025. PMID: 40362380 Free PMC article. Review.
References
-
- Aakalu G, Smith WB, Nguyen N, Jiang C, Schuman EM. Dynamic visualization of local protein synthesis in hippocampal neurons. Neuron. 2001;30:489–502. - PubMed
-
- Banko JL, Merhav M, Stern E, Sonenberg N, Rosenblum K, Klann E. Behavioral alterations in mice lacking the translation repressor 4E-BP2. Neurobiol Learn Mem. 2007;87:248–256. - PubMed
-
- Bekkers JM, Stevens CF. Cable properties of cultured hippocampal neurons determined from sucrose-evoked miniature EPSCs. J Neurophysiol. 1996;75:1250–1255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
