

Physical interaction of RECQ5 helicase with RAD51 facilitates its anti-recombinase activity
- PMID: 20348101
- PMCID: PMC2871440
- DOI: 10.1074/jbc.M110.110478
Physical interaction of RECQ5 helicase with RAD51 facilitates its anti-recombinase activity
Abstract
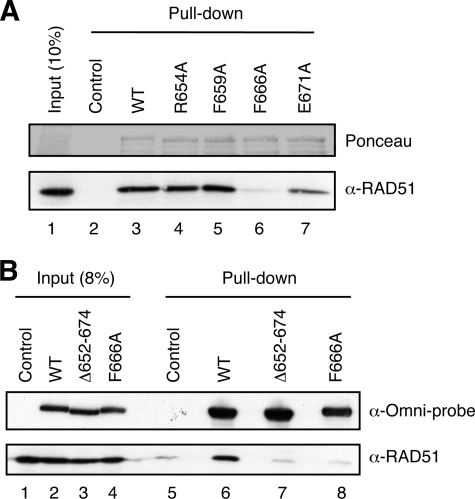
Homologous recombination (HR) provides an efficient mechanism for error-free repair of DNA double-strand breaks (DSBs). However, HR can be also harmful as inappropriate or untimely HR events can give rise to lethal recombination intermediates and chromosome rearrangements. A critical step of HR is the formation of a RAD51 filament on single-stranded (ss)DNA, which mediates the invasion of a homologous DNA molecule. In mammalian cells, several DNA helicases have been implicated in the regulation of this process. RECQ5, a member of the RecQ family of DNA helicases, interacts physically with the RAD51 recombinase and disrupts RAD51 presynaptic filaments in a reaction dependent on ATP hydrolysis. Here, we have precisely mapped the RAD51-interacting domain of RECQ5 and generated mutants that fail to interact with RAD51. We show that although these mutants retain normal ATPase activity, they are impaired in their ability to displace RAD51 from ssDNA. Moreover, we show that ablation of RECQ5-RAD51 complex formation by a point mutation alleviates the inhibitory effect of RECQ5 on HR-mediated DSB repair. These findings provide support for the proposal that interaction with RAD51 is critical for the anti-recombinase attribute of RECQ5.
Figures




Similar articles
-
Single-molecule visualization of human RECQ5 interactions with single-stranded DNA recombination intermediates.Nucleic Acids Res. 2021 Jan 11;49(1):285-305. doi: 10.1093/nar/gkaa1184. Nucleic Acids Res. 2021. PMID: 33332547 Free PMC article.
-
Human RECQ5 helicase promotes repair of DNA double-strand breaks by synthesis-dependent strand annealing.Nucleic Acids Res. 2014 Feb;42(4):2380-90. doi: 10.1093/nar/gkt1263. Epub 2013 Dec 6. Nucleic Acids Res. 2014. PMID: 24319145 Free PMC article.
-
Regulation of Rad51 recombinase presynaptic filament assembly via interactions with the Rad52 mediator and the Srs2 anti-recombinase.J Biol Chem. 2009 Sep 4;284(36):24363-71. doi: 10.1074/jbc.M109.032953. Epub 2009 Jul 15. J Biol Chem. 2009. PMID: 19605344 Free PMC article.
-
RecQ helicases queuing with Srs2 to disrupt Rad51 filaments and suppress recombination.Genes Dev. 2007 Dec 1;21(23):3019-26. doi: 10.1101/gad.1624707. Genes Dev. 2007. PMID: 18056418 Review. No abstract available.
-
Regulation of DNA strand exchange in homologous recombination.DNA Repair (Amst). 2010 Dec 10;9(12):1264-72. doi: 10.1016/j.dnarep.2010.09.014. DNA Repair (Amst). 2010. PMID: 20971042 Review.
Cited by
-
The MCM8-MCM9 complex promotes RAD51 recruitment at DNA damage sites to facilitate homologous recombination.Mol Cell Biol. 2013 Apr;33(8):1632-44. doi: 10.1128/MCB.01503-12. Epub 2013 Feb 11. Mol Cell Biol. 2013. PMID: 23401855 Free PMC article.
-
Structural mimicry in transcription regulation of human RNA polymerase II by the DNA helicase RECQL5.Nat Struct Mol Biol. 2013 Jul;20(7):892-9. doi: 10.1038/nsmb.2596. Epub 2013 Jun 9. Nat Struct Mol Biol. 2013. PMID: 23748380 Free PMC article.
-
Insights into the RecQ helicase mechanism revealed by the structure of the helicase domain of human RECQL5.Nucleic Acids Res. 2017 Apr 20;45(7):4231-4243. doi: 10.1093/nar/gkw1362. Nucleic Acids Res. 2017. PMID: 28100692 Free PMC article.
-
PARP3 is a promoter of chromosomal rearrangements and limits G4 DNA.Nat Commun. 2017 Apr 27;8:15110. doi: 10.1038/ncomms15110. Nat Commun. 2017. PMID: 28447610 Free PMC article.
-
Structural insights into transcriptional regulation by the helicase RECQL5.bioRxiv [Preprint]. 2025 Jan 29:2025.01.29.634372. doi: 10.1101/2025.01.29.634372. bioRxiv. 2025. Update in: Nat Struct Mol Biol. 2025 Jul 7. doi: 10.1038/s41594-025-01611-8. PMID: 39975028 Free PMC article. Updated. Preprint.
References
-
- San Filippo J., Sung P., Klein H. (2008) Annu. Rev. Biochem. 77, 229–257 - PubMed
-
- Sung P., Klein H. (2006) Nat. Rev. Mol. Cell Biol. 7, 739–750 - PubMed
-
- Gangloff S., Soustelle C., Fabre F. (2000) Nat. Genet. 25, 192–194 - PubMed
-
- Krejci L., Van Komen S., Li Y., Villemain J., Reddy M. S., Klein H., Ellenberger T., Sung P. (2003) Nature 423, 305–309 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
