

The Arabidopsis stem cell factor POLTERGEIST is membrane localized and phospholipid stimulated
- PMID: 20348433
- PMCID: PMC2861466
- DOI: 10.1105/tpc.109.068734
The Arabidopsis stem cell factor POLTERGEIST is membrane localized and phospholipid stimulated
Abstract
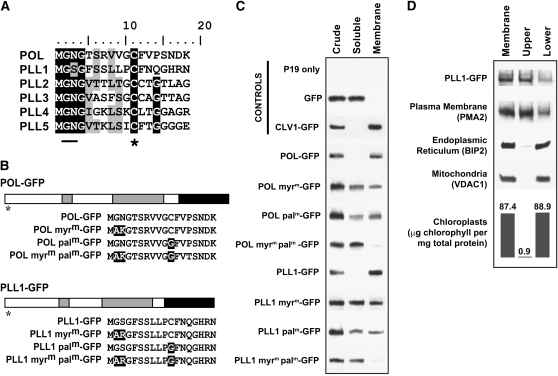
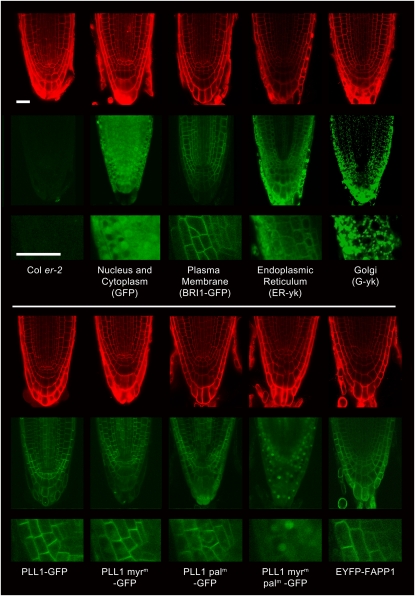
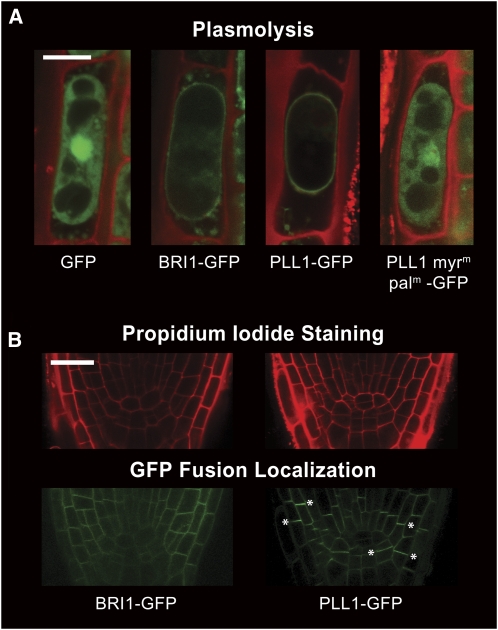
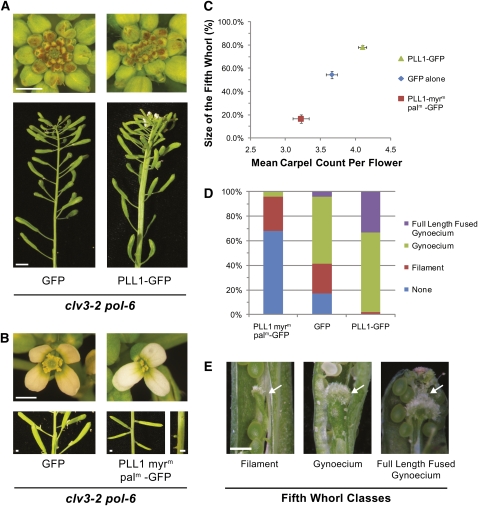
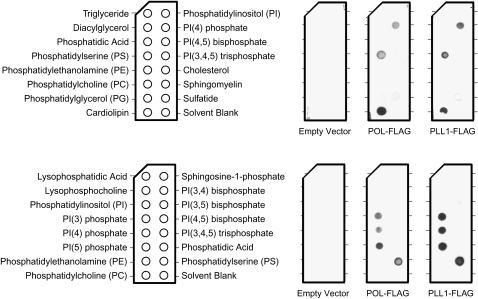
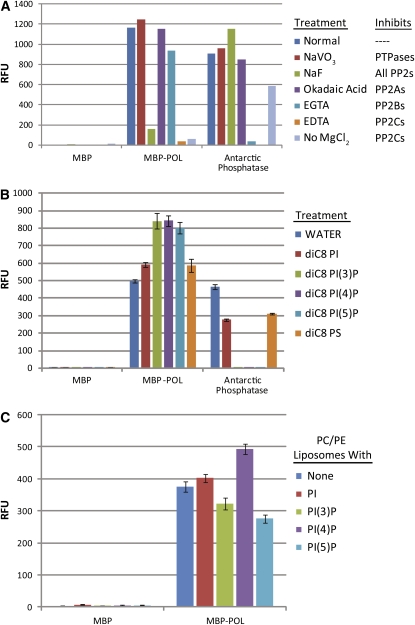
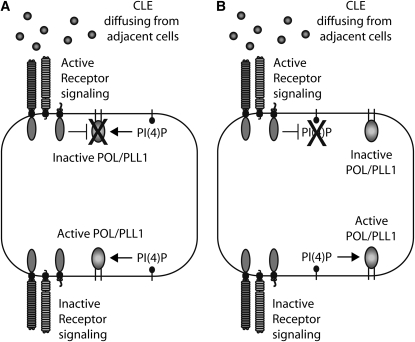
Stem cell maintenance and differentiation are tightly regulated in multicellular organisms. In plants, proper control of the stem cell populations is critical for extensive postembryonic organogenesis. The Arabidopsis thaliana protein phosphatase type 2C proteins POLTERGEIST (POL) and PLL1 are essential for maintenance of both the root and shoot stem cells. Specifically, POL and PLL1 are required for proper specification of key asymmetric cell divisions during stem cell initiation and maintenance. POL and PLL1 are known to be integral components of the CLE/WOX signaling pathways, but the location and mechanisms by which POL and PLL1 are regulated within these pathways are unclear. Here, we show that POL and PLL1 are dual-acylated plasma membrane proteins whose membrane localization is required for proper function. Furthermore, this localization places POL and PLL1 in proximity of the upstream plasma membrane receptors that regulate their activity. Additionally, we find that POL and PLL1 directly bind to multiple lipids and that POL is catalytically activated by phosphatidylinositol (4) phosphate [PI(4)P] in vitro. Based on these results, we propose that the upstream receptors in the CLE/WOX signaling pathways may function to either limit PI(4)P availability or antagonize PI(4)P stimulation of POL/PLL1. Significantly, the findings presented here suggest that phospholipids play an important role in promoting stem cell specification.
Figures
References
-
- Alvarez-Venegas R., Sadder M., Hlavacka A., Baluska F., Xia Y., Lu G., Firsov A., Sarath G., Moriyama H., Dubrovsky J.G., Avramova Z. (2006). The Arabidopsis homolog of trithorax, ATX1, binds phosphatidylinositol 5-phosphate, and the two regulate a common set of target genes. Proc. Natl. Acad. Sci. USA 103: 6049–6054 - PMC - PubMed
-
- Baudouin E., Meskiene I., Hirt H. (1999). Short communication: Unsaturated fatty acids inhibit MP2C, a protein phosphatase 2C involved in the wound-induced MAP kinase pathway regulation. Plant J. 20: 343–348 - PubMed
-
- Boisson B., Giglione C., Meinnel T. (2003). Unexpected protein families including cell defense components feature in the N-myristoylome of a higher eukaryote. J. Biol. Chem. 278: 43418–43429 - PubMed
-
- Brand U., Fletcher J.C., Hobe M., Meyerowitz E.M., Simon R. (2000). Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 289: 617–619 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
