

Reduction in endocannabinoid tone is a homeostatic mechanism for specific inhibitory synapses
- PMID: 20348918
- PMCID: PMC2860695
- DOI: 10.1038/nn.2517
Reduction in endocannabinoid tone is a homeostatic mechanism for specific inhibitory synapses
Abstract
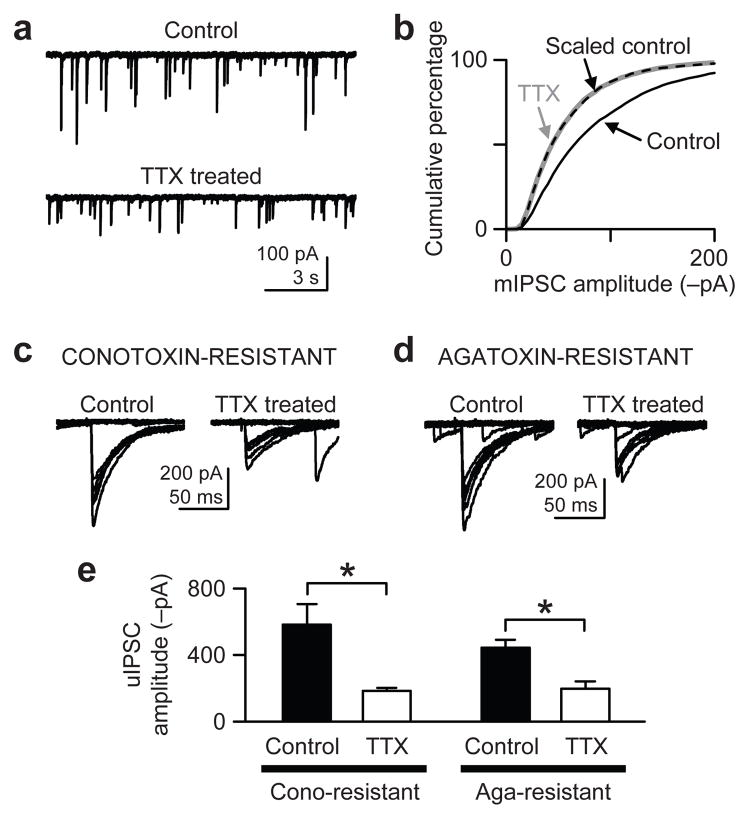
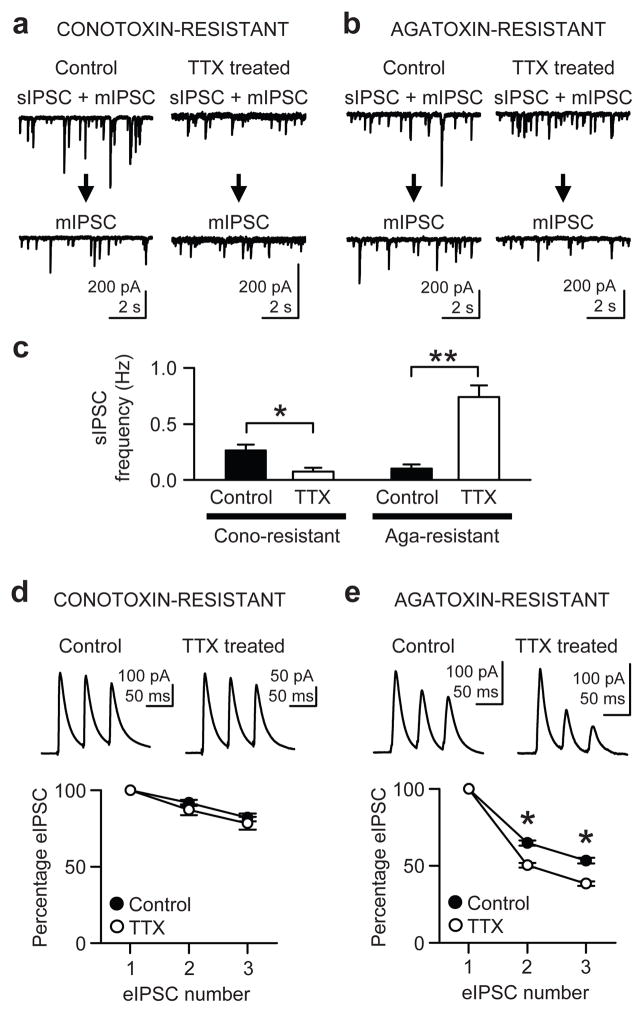
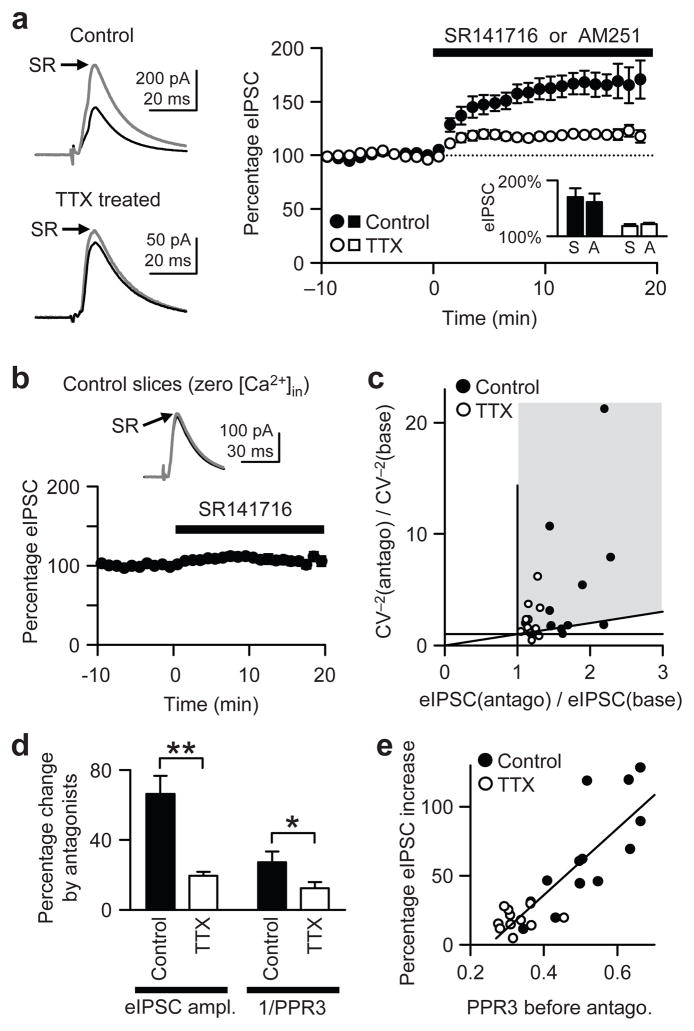
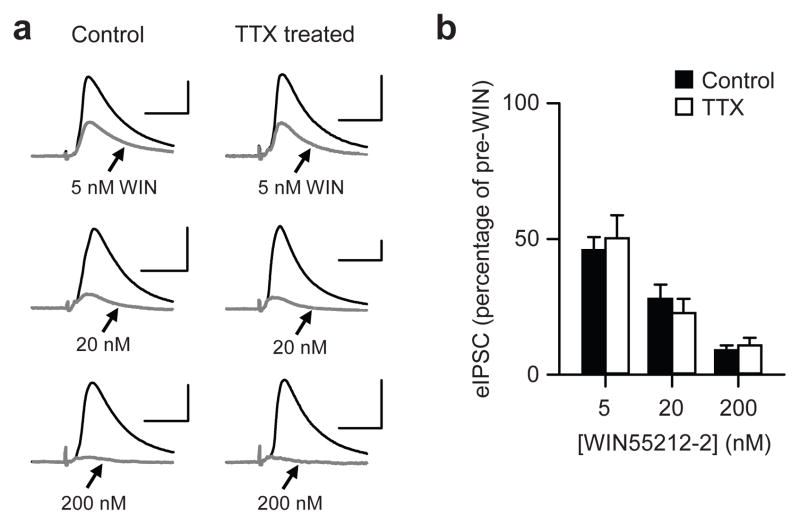
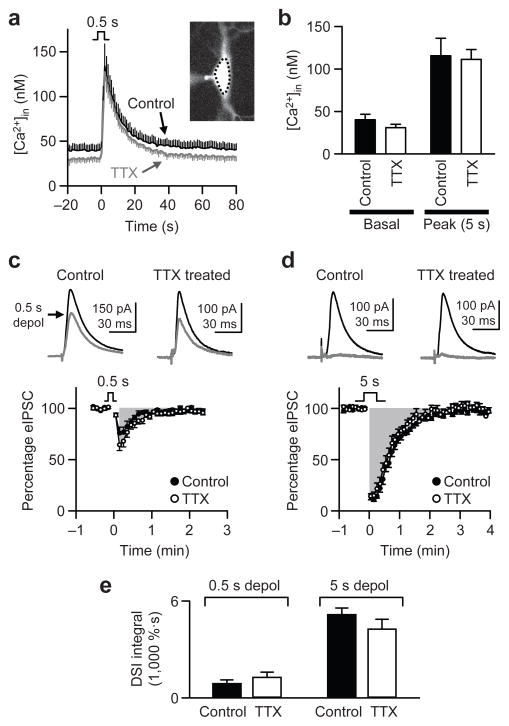
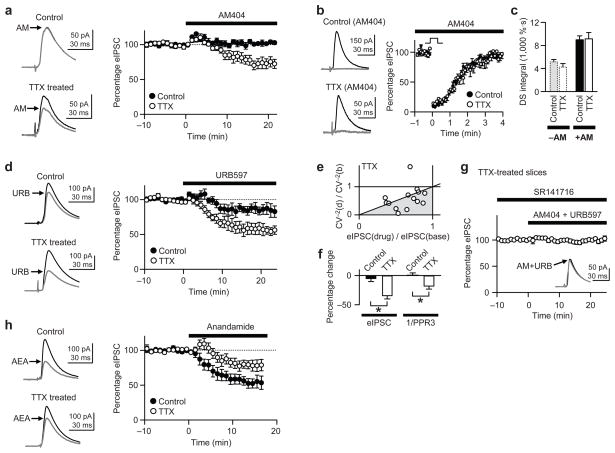
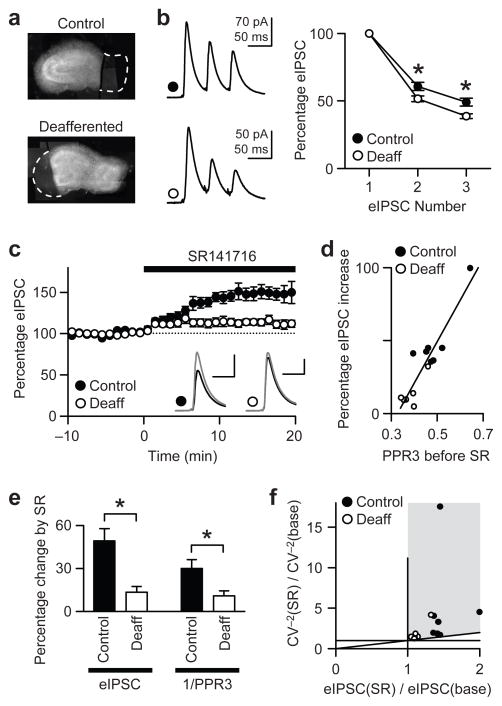
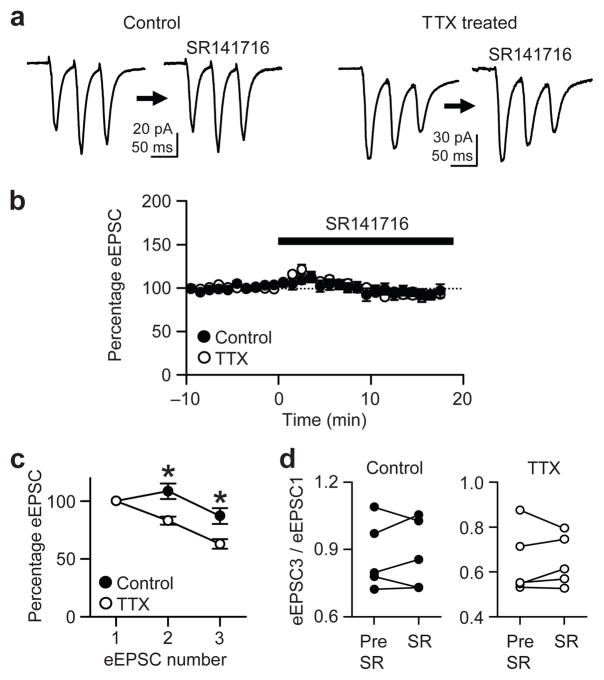
When chronic alterations in neuronal activity occur, network gain is maintained by global homeostatic scaling of synaptic strength, but the stability of microcircuits can be controlled by unique adaptations that differ from the global changes. It is not understood how specificity of synaptic tuning is achieved. We found that, although a large population of inhibitory synapses was homeostatically scaled down after chronic inactivity, decreased endocannabinoid tone specifically strengthened a subset of GABAergic synapses that express cannabinoid receptors. In rat hippocampal slice cultures, a 3-5-d blockade of neuronal firing facilitated uptake and degradation of anandamide. The consequent reduction in basal stimulation of cannabinoid receptors augmented GABA release probability, fostering rapid depression of synaptic inhibition and on-demand disinhibition. This regulatory mechanism, mediated by activity-dependent changes in tonic endocannabinoid level, permits selective local tuning of inhibitory synapses in hippocampal networks.
Figures
Similar articles
-
Endocannabinoid- and mGluR5-dependent short-term synaptic depression in an isolated neuron/bouton preparation from the hippocampal CA1 region.J Neurophysiol. 2008 Aug;100(2):1041-52. doi: 10.1152/jn.90226.2008. Epub 2008 May 21. J Neurophysiol. 2008. PMID: 18497350 Free PMC article.
-
Depolarizing GABAergic synaptic input triggers endocannabinoid-mediated retrograde synaptic signaling.Synapse. 2009 Aug;63(8):643-52. doi: 10.1002/syn.20641. Synapse. 2009. PMID: 19347961
-
Retrograde endocannabinoid signaling at striatal synapses requires a regulated postsynaptic release step.Proc Natl Acad Sci U S A. 2007 Dec 18;104(51):20564-9. doi: 10.1073/pnas.0706873104. Epub 2007 Dec 11. Proc Natl Acad Sci U S A. 2007. PMID: 18077376 Free PMC article.
-
A synaptogenic amide N-docosahexaenoylethanolamide promotes hippocampal development.Prostaglandins Other Lipid Mediat. 2011 Nov;96(1-4):114-20. doi: 10.1016/j.prostaglandins.2011.07.002. Epub 2011 Jul 23. Prostaglandins Other Lipid Mediat. 2011. PMID: 21810478 Free PMC article. Review.
-
The self-tuning neuron: synaptic scaling of excitatory synapses.Cell. 2008 Oct 31;135(3):422-35. doi: 10.1016/j.cell.2008.10.008. Cell. 2008. PMID: 18984155 Free PMC article. Review.
Cited by
-
Coordinated Regulation of CB1 Cannabinoid Receptors and Anandamide Metabolism Stabilizes Network Activity during Homeostatic Downscaling.eNeuro. 2022 Nov 11;9(6):ENEURO.0276-22.2022. doi: 10.1523/ENEURO.0276-22.2022. Print 2022 Nov-Dec. eNeuro. 2022. PMID: 36316118 Free PMC article.
-
Electrophysiology of Endocannabinoid Signaling.Methods Mol Biol. 2023;2576:461-475. doi: 10.1007/978-1-0716-2728-0_38. Methods Mol Biol. 2023. PMID: 36152210
-
Tempering aversive/traumatic memories with cannabinoids: a review of evidence from animal and human studies.Psychopharmacology (Berl). 2019 Jan;236(1):201-226. doi: 10.1007/s00213-018-5127-x. Epub 2019 Jan 2. Psychopharmacology (Berl). 2019. PMID: 30604182 Review.
-
Activity-dependent adaptations in inhibitory axons.Front Cell Neurosci. 2013 Nov 21;7:219. doi: 10.3389/fncel.2013.00219. Front Cell Neurosci. 2013. PMID: 24312009 Free PMC article. Review.
-
Illuminating the multifaceted roles of neurotransmission in shaping neuronal circuitry.Neuron. 2014 Sep 17;83(6):1303-1318. doi: 10.1016/j.neuron.2014.08.029. Neuron. 2014. PMID: 25233313 Free PMC article. Review.
References
-
- Trasande CA, Ramirez JM. Activity deprivation leads to seizures in hippocampal slice cultures: is epilepsy the consequence of homeostatic plasticity? J Clin Neurophysiol. 2007;24:154–164. - PubMed
-
- Bausch SB, He S, Petrova Y, Wang XM, McNamara JO. Plasticity of both excitatory and inhibitory synapses is associated with seizures induced by removal of chronic blockade of activity in cultured hippocampus. J Neurophysiol. 2006;96:2151–2167. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
