

The proapoptotic BH3-only protein Bim is downregulated in a subset of colorectal cancers and is repressed by antiapoptotic COX-2/PGE(2) signalling in colorectal adenoma cells
- PMID: 20348947
- PMCID: PMC2883743
- DOI: 10.1038/onc.2010.94
The proapoptotic BH3-only protein Bim is downregulated in a subset of colorectal cancers and is repressed by antiapoptotic COX-2/PGE(2) signalling in colorectal adenoma cells
Abstract
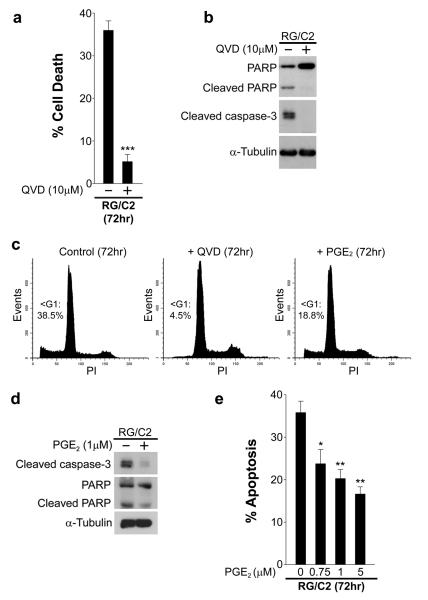
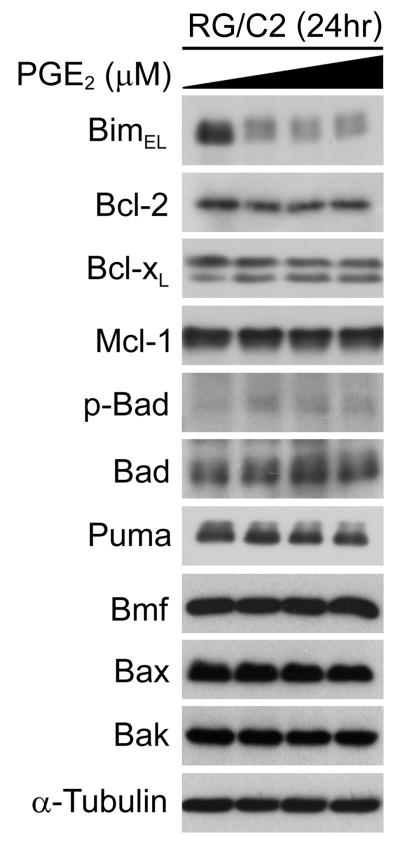
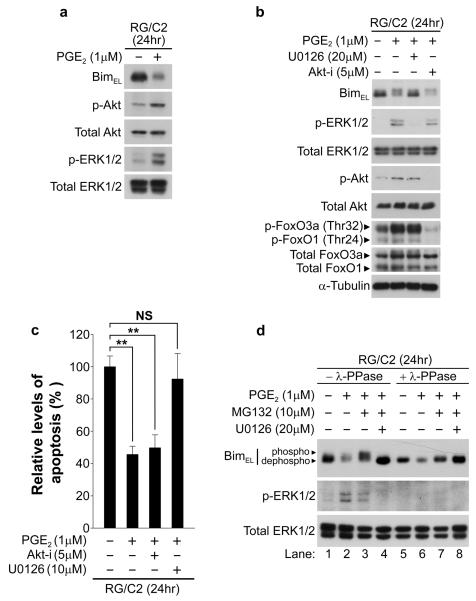
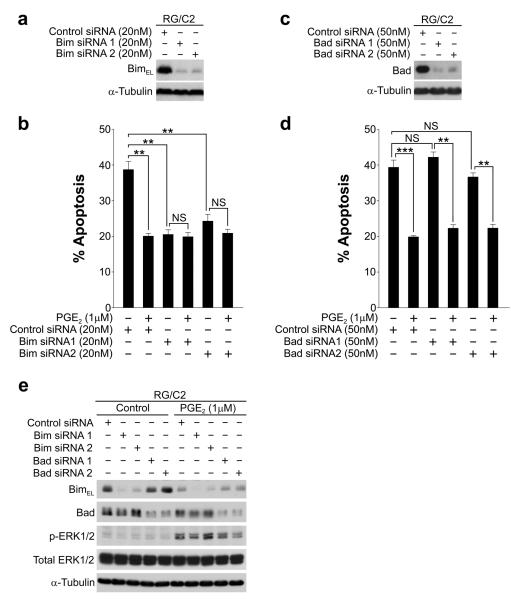
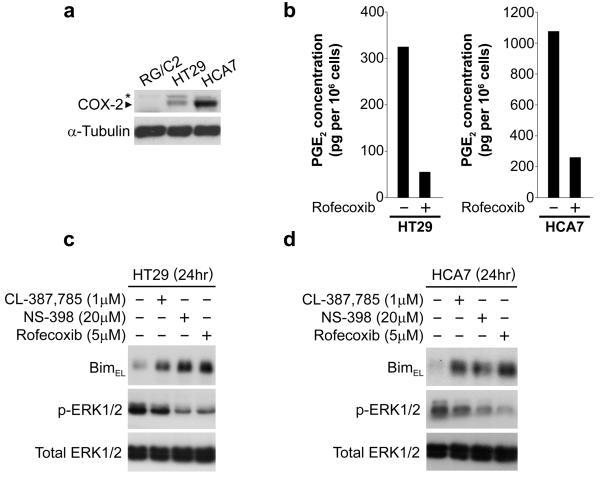
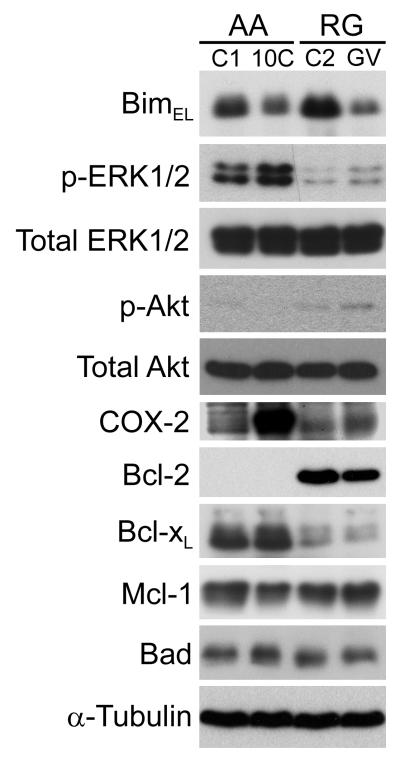
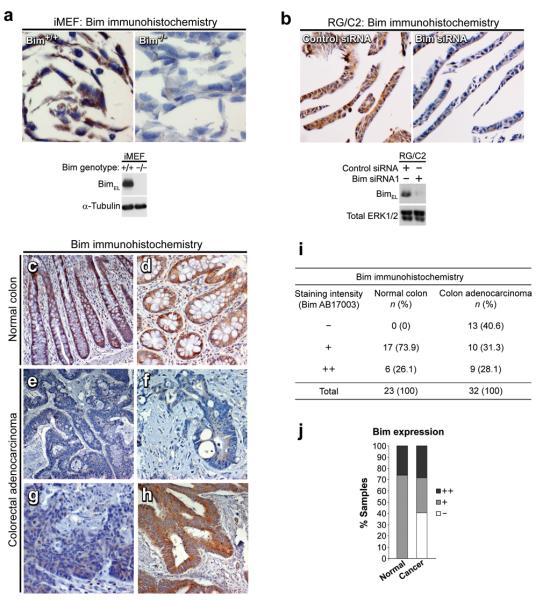
Overexpression of cyclooxygenase-2 (COX-2) and elevated levels of its enzymatic product prostaglandin E2 (PGE(2)) occur in the majority of colorectal cancers and have important roles in colorectal tumorigenesis. However, despite the established prosurvival role of PGE(2) in cancer, the underlying mechanisms are not fully understood. Here, we have shown that PGE(2) suppresses apoptosis via repression of the proapoptotic BH3-only protein Bim in human colorectal adenoma cells. Repression of Bim expression was dependent upon PGE(2)-mediated activation of the Raf-MEK-ERK1/2 pathway, which promoted Bim phosphorylation and proteasomal degradation. Reduction of Bim expression using RNA interference reduced spontaneous apoptosis in adenoma cells and abrogated PGE(2)-dependent apoptosis suppression. Treatment of COX-2-expressing colorectal carcinoma cells with COX-2-selective NSAIDs-induced Bim expression, suggesting that Bim repression via PGE(2) signalling may be opposed by COX-2 inhibition. Examination of Bim expression in two established in vitro models of the adenoma-carcinoma sequence revealed that downregulation of Bim expression was associated with tumour progression towards an anchorage-independent phenotype. Finally, immunohistochemical analyses revealed that Bim expression is markedly reduced in approximately 40% of human colorectal carcinomas in vivo. These observations highlight the COX-2/PGE(2) pathway as an important negative regulator of Bim expression in colorectal tumours and suggest that Bim repression may be an important step during colorectal cancer tumorigenesis.
Figures
References
-
- Balmanno K, Cook SJ. Tumour cell survival signalling by the ERK1/2 pathway. Cell Death Differ. 2009;16:368–77. - PubMed
-
- Bedi A, Pasricha PJ, Akhtar AJ, Barber JP, Bedi GC, Giardiello FM, et al. Inhibition of apoptosis during development of colorectal cancer. Cancer Res. 1995;55:1811–6. - PubMed
-
- Bouillet P, Metcalf D, Huang DC, Tarlinton DM, Kay TW, Kontgen F, et al. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. Science. 1999;286:1735–8. - PubMed
-
- Bresalier RS, Sandler RS, Quan H, Bolognese JA, Oxenius B, Horgan K, et al. Cardiovascular events associated with rofecoxib in a colorectal adenoma chemoprevention trial. N Engl J Med. 2005;352:1092–102. - PubMed
-
- Brown JR, DuBois RN. COX-2: a molecular target for colorectal cancer prevention. J Clin Oncol. 2005;23:2840–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
