

SSBP2 is an in vivo tumor suppressor and regulator of LDB1 stability
- PMID: 20348955
- PMCID: PMC2878399
- DOI: 10.1038/onc.2010.78
SSBP2 is an in vivo tumor suppressor and regulator of LDB1 stability
Abstract
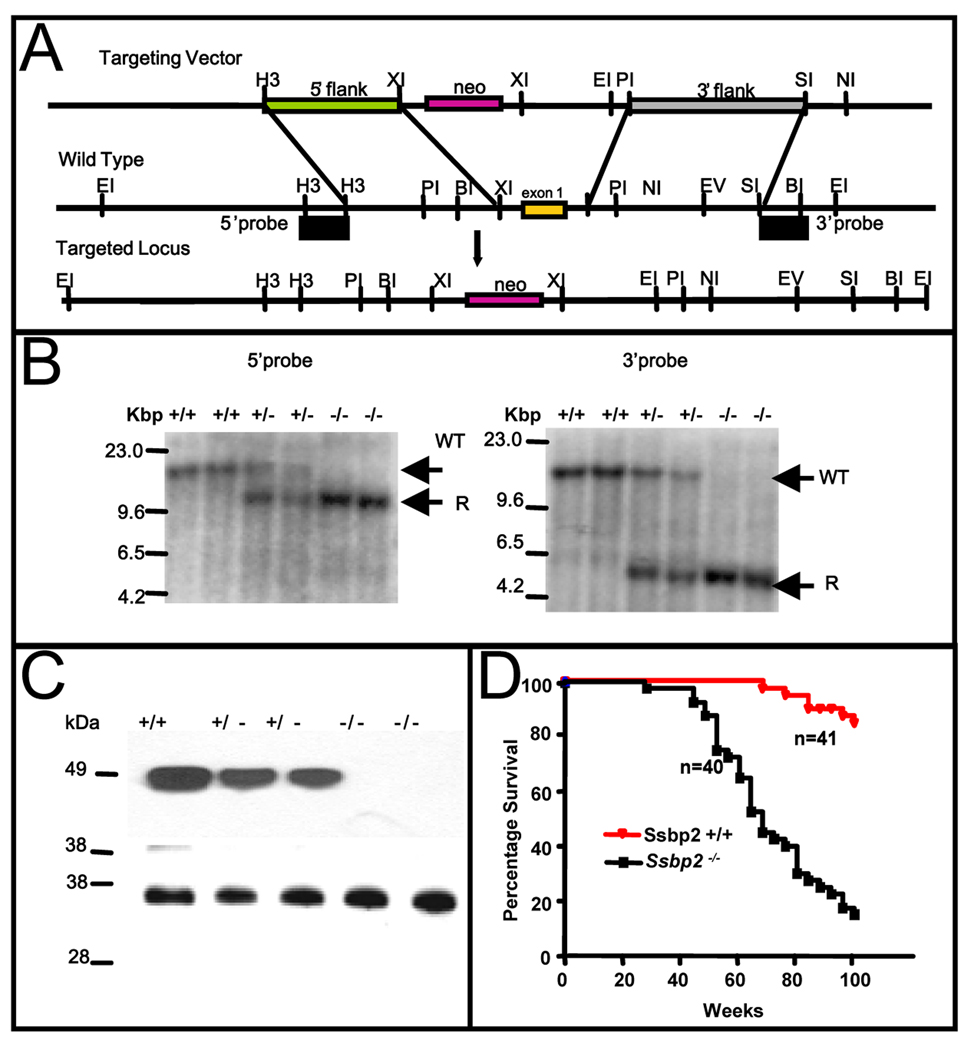
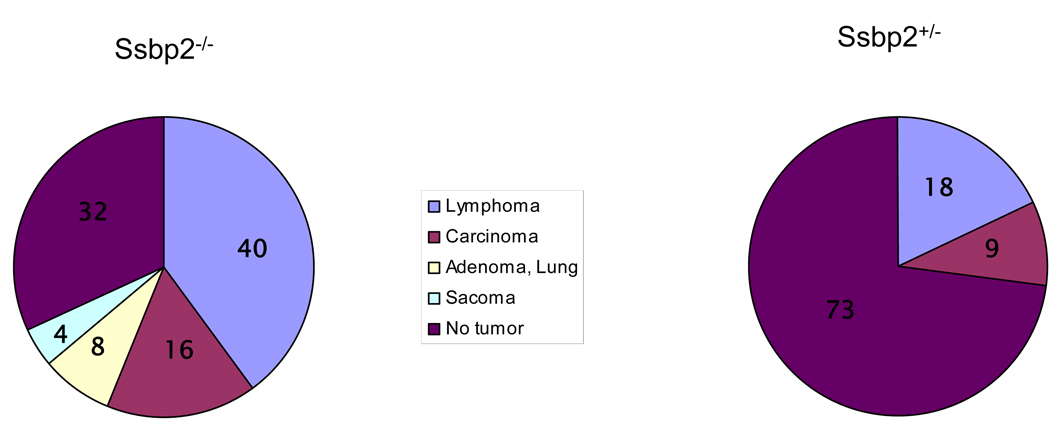
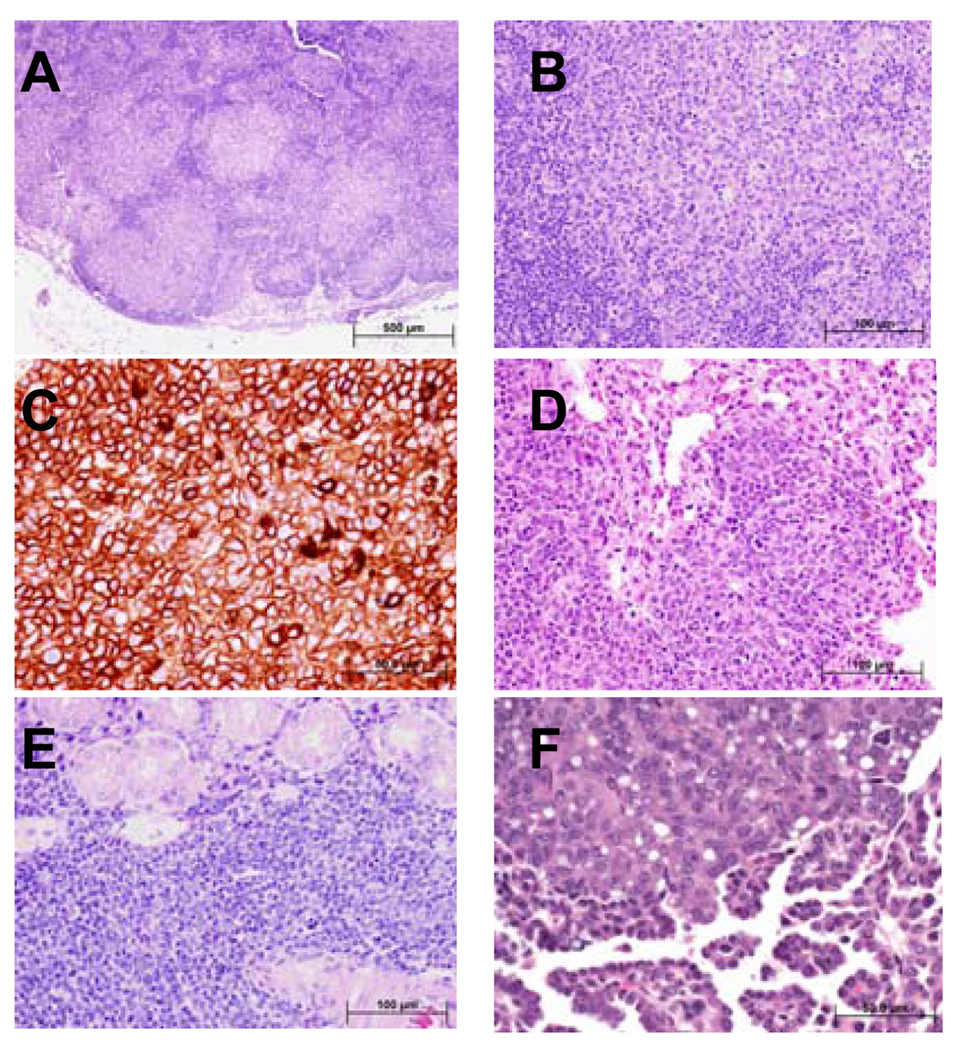
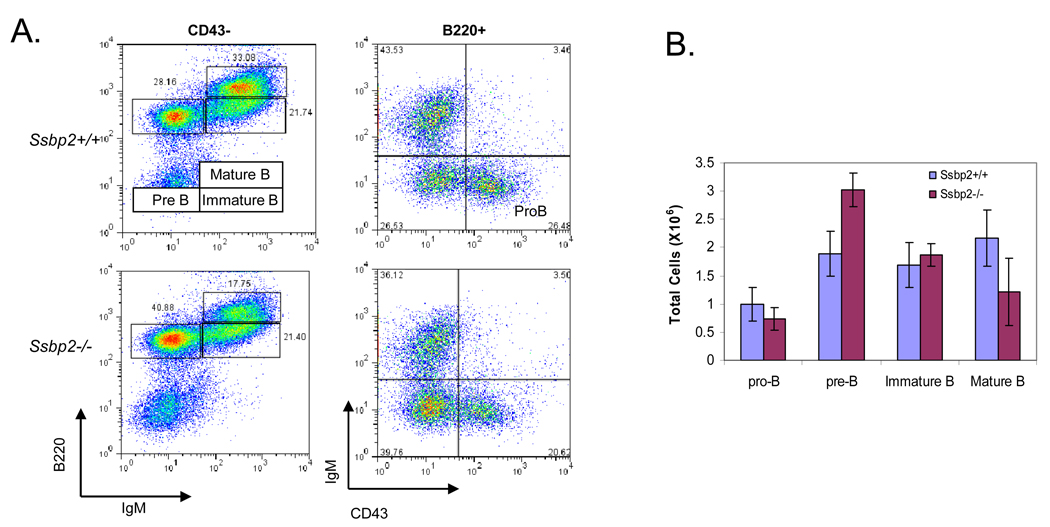
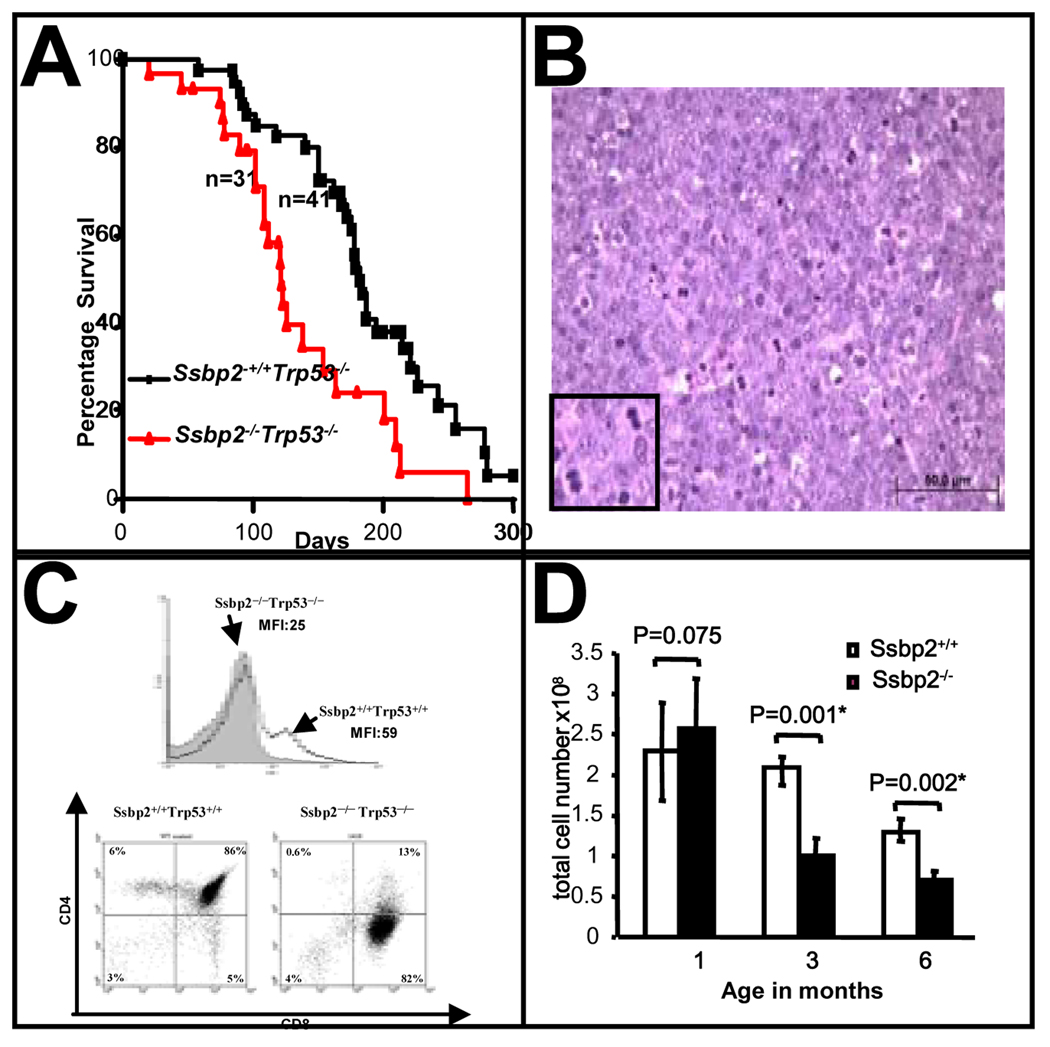
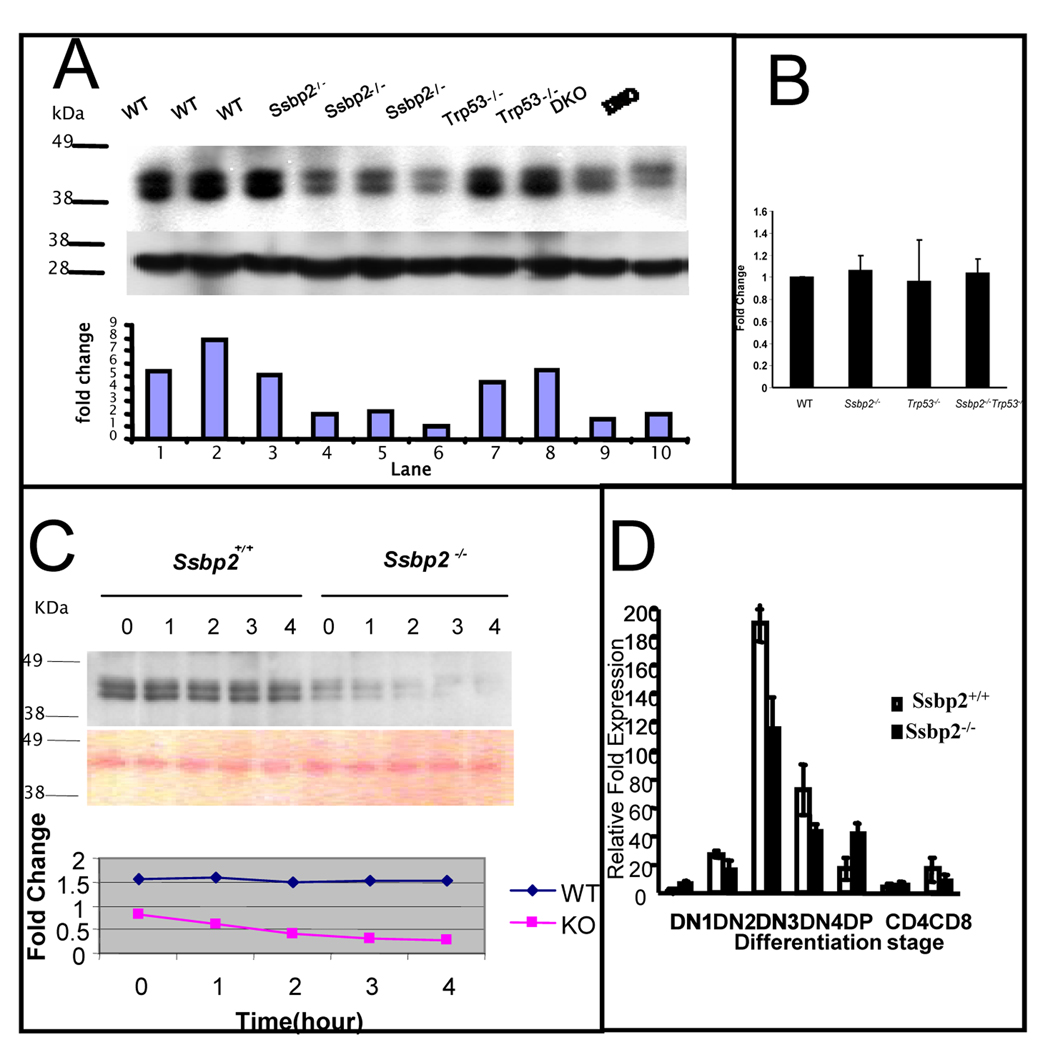
SSBP proteins bind and stabilize transcriptional cofactor LIM domain-binding protein1 (LDB1) from proteosomal degradation to promote tissue-specific transcription through an evolutionarily conserved pathway. The human SSBP2 gene was isolated as a candidate tumor suppressor from a critical region of loss in chromosome 5q14.1. By gene targeting, we show increased predisposition to B-cell lymphomas and carcinomas in Ssbp2(-/-) mice. Remarkably, loss of Ssbp2 causes increased LDB1 turnover in the thymus, a pathway exploited in Trp53(-/-)Ssbp2(-/-) mice to develop highly aggressive, immature thymic lymphomas. Using T-cell differentiation as a model, we report a stage-specific upregulation of Ssbp2 expression, which in turn regulates LDB1 turnover under physiological conditions. Furthermore, transcript levels of pTalpha, a target of LDB1-containing complex, and a critical regulator T-cell differentiation are reduced in Ssbp2(-/-) immature thymocytes. Our findings suggest that disruption of the SSBP2-regulated pathways may be an infrequent but critical step in malignant transformation of multiple tissues.
Conflict of interest statement
The authors do not have any competing financial interest in the results presented.
Figures
References
-
- Agulnick AD, Taira M, Breen JJ, Tanaka T, Dawid IB, Westphal H. Nature. 1996;384:270–272. - PubMed
-
- Aoyama M, Ozaki T, Inuzuka H, Tomotsune D, Hirato J, Okamoto Y, Tokita H, Ohira M, Nakagawara A. Cancer Res. 2005;65:4587–4597. - PubMed
-
- Bach I, Rodriguez-Esteban C, Carriere C, Bhushan A, Krones A, Rose DW, Glass CK, Andersen B, Izpisua Belmonte JC, Rosenfeld MG. Nat Genet. 1999;22:394–399. - PubMed
-
- Baseta JG, Stutman O. J Immunol. 2000;165:5621–5630. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
