

Critical determinants for chromatin binding by Saccharomyces cerevisiae Yng1 exist outside of the plant homeodomain finger
- PMID: 20351216
- PMCID: PMC2881130
- DOI: 10.1534/genetics.110.116285
Critical determinants for chromatin binding by Saccharomyces cerevisiae Yng1 exist outside of the plant homeodomain finger
Abstract
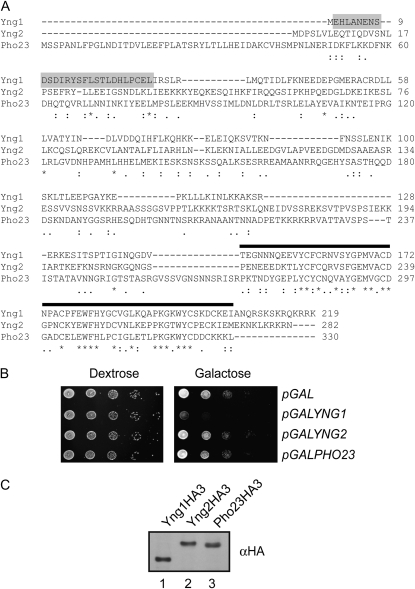
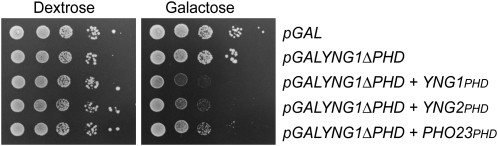
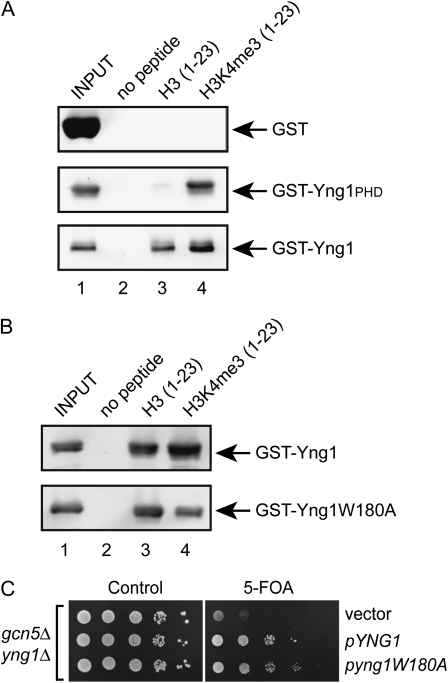
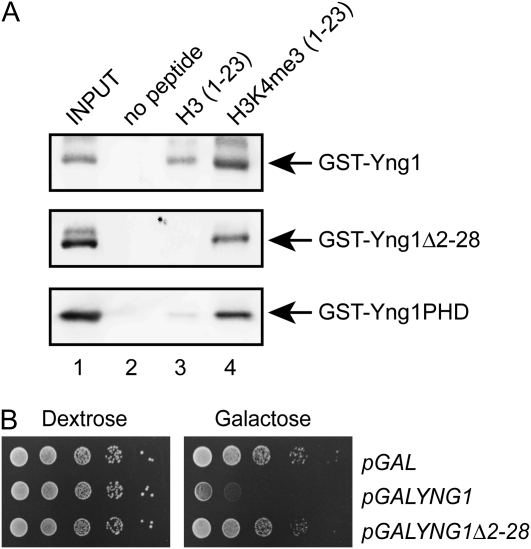
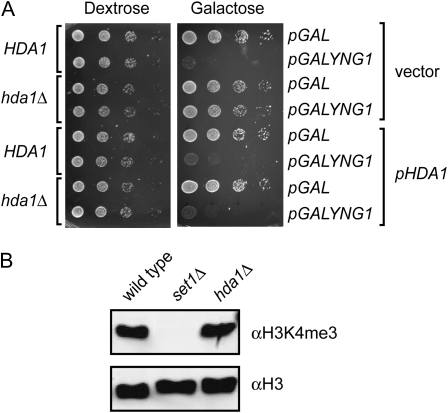
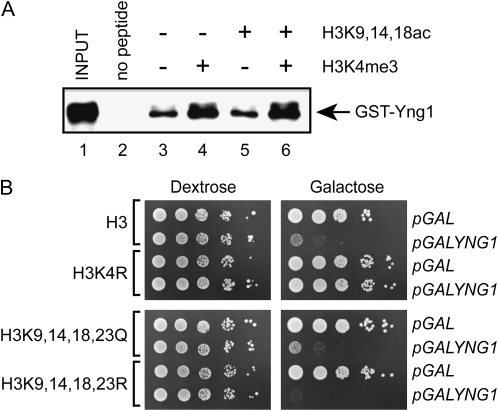
The temporal and spatial regulation of histone post-translational modifications is essential for proper chromatin structure and function. The Saccharomyces cerevisiae NuA3 histone acetyltransferase complex modifies the amino-terminal tail of histone H3, but how NuA3 is targeted to specific regions of the genome is not fully understood. Yng1, a subunit of NuA3 and a member of the Inhibitor of Growth (ING) protein family, is required for the interaction of NuA3 with chromatin. This protein contains a C-terminal plant homeodomain (PHD) finger that specifically interacts with lysine 4-trimethylated histone H3 (H3K4me3) in vitro. This initially suggested that NuA3 is targeted to regions bearing the H3K4me3 mark; however, deletion of the Yng1 PHD finger does not disrupt the interaction of NuA3 with chromatin or result in a phenotype consistent with loss of NuA3 function in vivo. In this study, we uncovered the molecular basis for the discrepancies in these data. We present both genetic and biochemical evidence that full-length Yng1 has two independent histone-binding motifs: an amino-terminal motif that binds unmodified H3 tails and a carboxyl-terminal PHD finger that specifically recognizes H3K4me3. Although these motifs can bind histones independently, together they increase the apparent association of Yng1 for the H3 tail.
Figures
References
-
- Albert, I., T. N. Mavrich, L. P. Tomsho, J. Qi, S. J. Zanton et al., 2007. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446 572–576. - PubMed
-
- Ausubel, F. M., 1987. Current Protocols in Molecular Biology. Greene Publishing Associates and Wiley-Interscience/John Wiley & Sons, New York.
-
- Carmen, A. A., S. E. Rundlett and M. Grunstein, 1996. HDA1 and HDA3 are components of a yeast histone deacetylase (HDA) complex. J. Biol. Chem. 271 15837–15844. - PubMed
-
- Carrozza, M. J., B. Li, L. Florens, T. Suganuma, S. K. Swanson et al., 2005. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell 123 581–592. - PubMed
-
- Choy, J. S., B. T. Tobe, J. H. Huh and S. J. Kron, 2001. Yng2p-dependent NuA4 histone H4 acetylation activity is required for mitotic and meiotic progression. J. Biol. Chem. 276 43653–43662. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
