

Review
doi: 10.1186/gb-2010-11-3-204.
Epub 2010 Mar 29.
The yin and yang of chromatin spatial organization
Affiliations
- PMID: 20353545
- PMCID: PMC2864563
- DOI: 10.1186/gb-2010-11-3-204
Item in Clipboard
Review
The yin and yang of chromatin spatial organization
Genome Biol.
2010.
Abstract
Spatial organization of the genome is non-random. Preferential chromatin interactions, both in cis and in trans and between transcriptionally active and silent regions, influence organization.
Figures
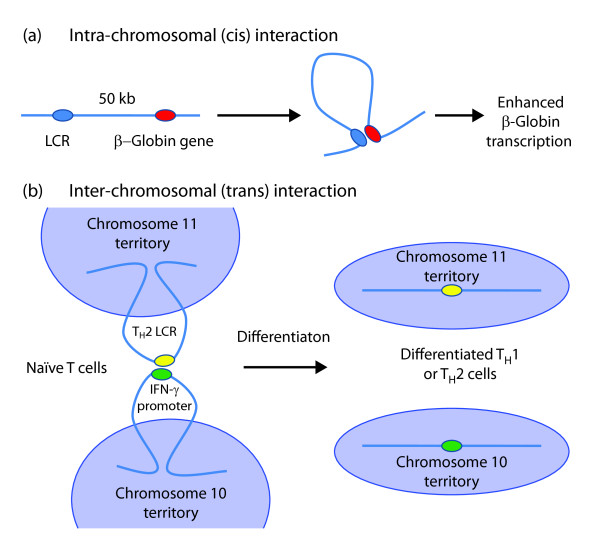
Intra- and inter-chromosomal interactions. (a) The β-globin gene, located approximately 50 kb downstream of the locus control region (LCR), is activated during erythropoiesis. The β-globin gene interaction with the LCR ensures high and efficient β-globin transcription, with the intervening sequence looping out. (b) Naïve T cells show a trans association between the TH2 LCR, on chromosome 11, and the IFN-γ promoter, on chromosome 10. This interaction is lost in favor of specific intra-chromosomal interactions following differentiation into TH1 or TH2 effector cells.
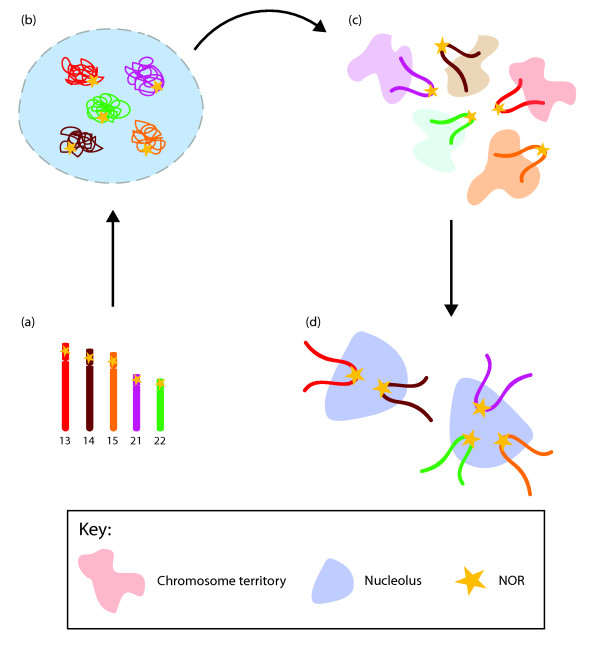
NORs cluster as cells exit mitosis. (a) The short arms of acrocentric chromosomes 13, 14, 15, 21 and 22 contain NORs, which are separated during mitosis. (b) As cells exit mitosis and the nuclear membrane begins to reform, chromosomes begin to decondense. (c) Loops of chromatin may extend away from the core of the territory. (d) As G1 phase is established and nucleoli form, loops of NOR-containing chromatin co-associate with the other components of the nucleolus and ribosomal DNA gene transcription is initiated.
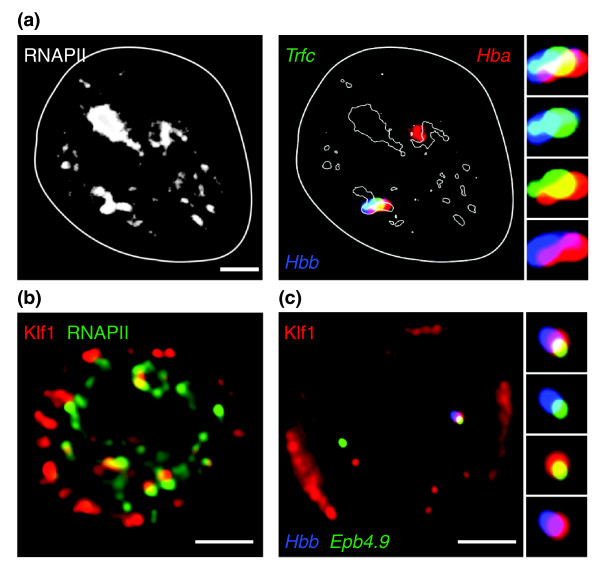
Colocalization of like-regulated genes and specialized transcription factories. (a) Quadruple-label RNA immuno-FISH of three genes that are being transcribed and their association with RNAPII transcription factories. RNAPII staining is shown on the left and an overlay of the RNAPII staining showing the contributions of the genes is on the right. The side panels show the enlarged images of colocalizing FISH signals, showing that transcription factories can simultaneously transcribe at least three genes, located on different chromosomes. (b) Immunofluorescence detection of Klf1 (red) and RNAPII transcription factories (green), showing the selective and specialized nature of transcription factories. (c) Triple-label RNA immuno-FISH for Hbb and Epb4.9, showing association of these genes at Klf1 foci. All images show definitive erythroid cells and the scale bar in each panel represents 2 μm. Reproduced, with permission, from [24].
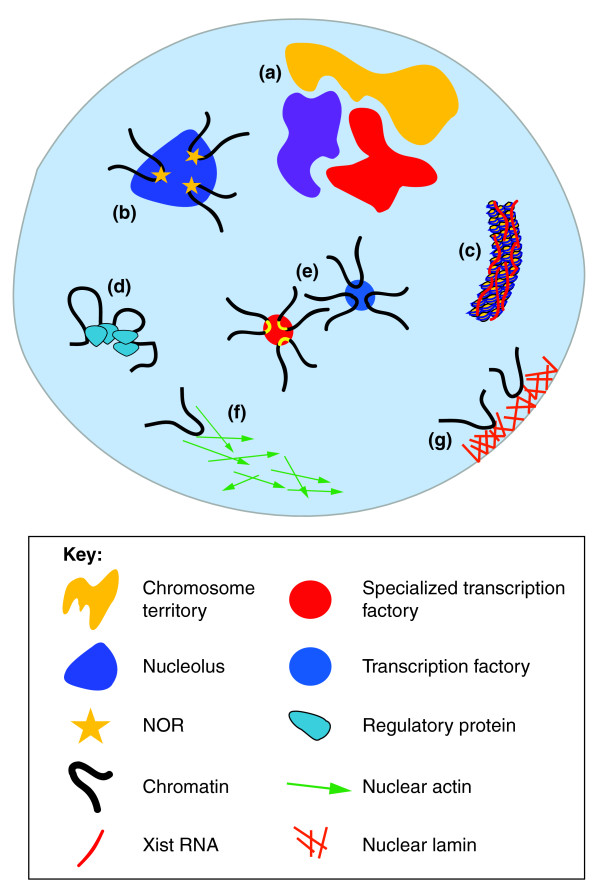
Schematic summary of some of the processes and structures that influence the spatial organization of the genome. Although not exhaustive, the figure depicts: (a) chromosome territories; (b) nucleoli and genomic regions clustering through nucleolar organizing regions (NOR); (c) the X chromosome and Xist RNA; (d) regulatory proteins such as CTCF, transcription factors and Polycomb repressive complexes (PRCs) that can induce loops between genomic elements; (e) transcription factories (blue) and specialized transcription factories (red); (f) the potential role of nuclear actin in mediating long-range chromatin movement; and (g) the interactions of chromatin regions with the nuclear lamina. These processes, along with others described in this article and many more, are likely be important in dynamically shaping the spatial environment and organization of the genome.
References
-
- Starck J, Sarkar R, Romana M, Bhargava A, Scarpa AL, Tanaka M, Chamberlain JW, Weissman SM, Forget BG. Developmental regulation of human gamma-globin and beta-globin genes in the absence of the locus-control region. Blood. 1994;84:1656–1665. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/E017460/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/B/0000C151/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/B/0000M723/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0800036/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
