

The human Cdc34 carboxyl terminus contains a non-covalent ubiquitin binding activity that contributes to SCF-dependent ubiquitination
- PMID: 20353940
- PMCID: PMC2878539
- DOI: 10.1074/jbc.M109.090621
The human Cdc34 carboxyl terminus contains a non-covalent ubiquitin binding activity that contributes to SCF-dependent ubiquitination
Abstract
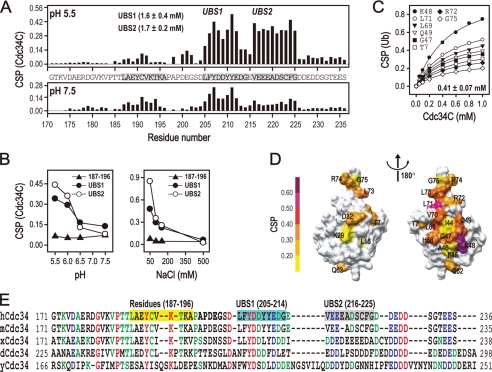
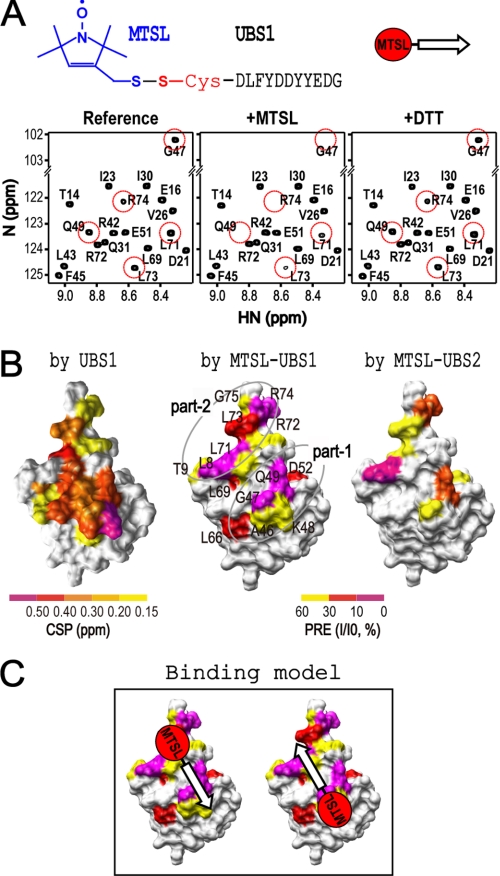
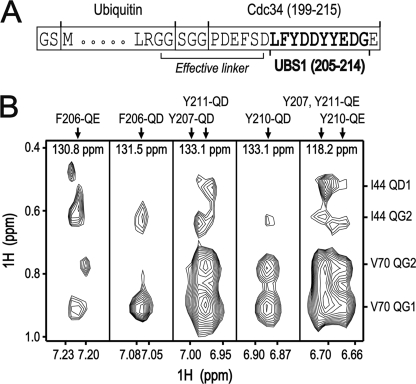
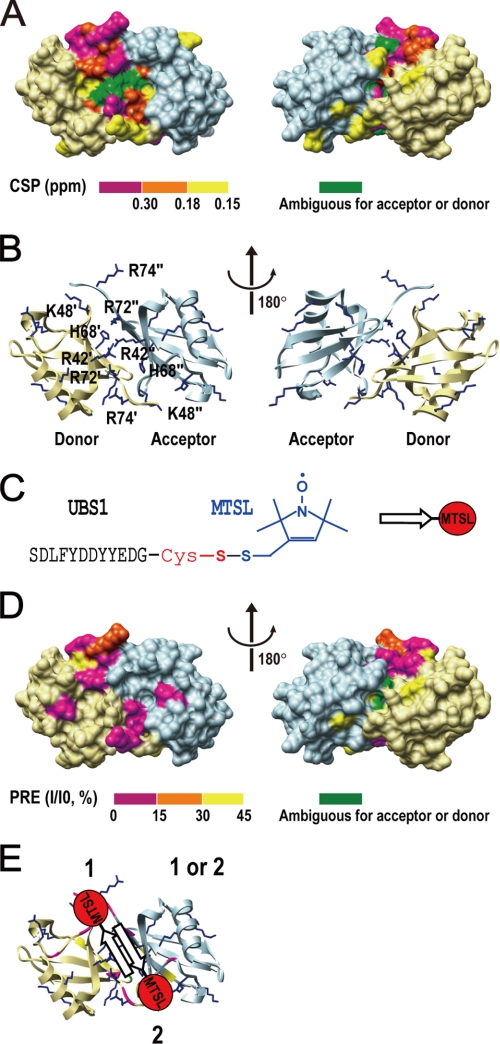
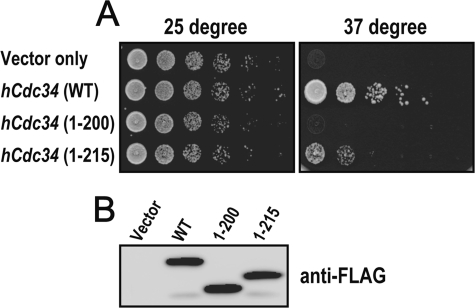
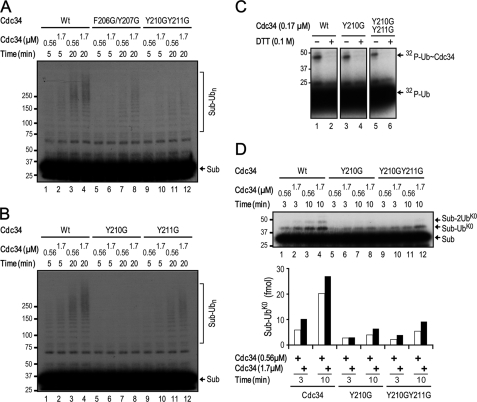
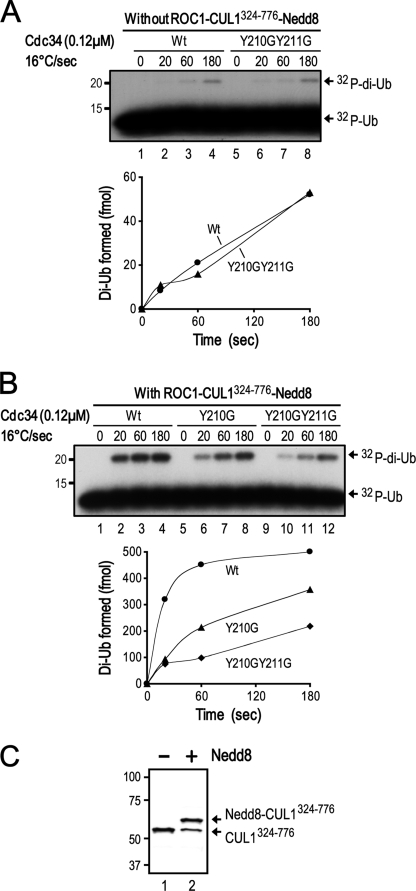
Cdc34 is an E2 ubiquitin-conjugating enzyme that functions in conjunction with SCF (Skp1.Cullin 1.F-box) E3 ubiquitin ligase to catalyze covalent attachment of polyubiquitin chains to a target protein. Here we identified direct interactions between the human Cdc34 C terminus and ubiquitin using NMR chemical shift perturbation assays. The ubiquitin binding activity was mapped to two separate Cdc34 C-terminal motifs (UBS1 and UBS2) that comprise residues 206-215 and 216-225, respectively. UBS1 and UBS2 bind to ubiquitin in the proximity of ubiquitin Lys(48) and C-terminal tail, both of which are key sites for conjugation. When bound to ubiquitin in one orientation, the Cdc34 UBS1 aromatic residues (Phe(206), Tyr(207), Tyr(210), and Tyr(211)) are probably positioned in the vicinity of ubiquitin C-terminal residue Val(70). Replacement of UBS1 aromatic residues by glycine or of ubiquitin Val(70) by alanine decreased UBS1-ubiquitin affinity interactions. UBS1 appeared to support the function of Cdc34 in vivo because human Cdc34(1-215) but not Cdc34(1-200) was able to complement the growth defect by yeast Cdc34 mutant strain. Finally, reconstituted IkappaBalpha ubiquitination analysis revealed a role for each adjacent pair of UBS1 aromatic residues (Phe(206)/Tyr(207), Tyr(210)/Tyr(211)) in conjugation, with Tyr(210) exhibiting the most pronounced catalytic function. Intriguingly, Cdc34 Tyr(210) was required for the transfer of the donor ubiquitin to a receptor lysine on either IkappaBalpha or a ubiquitin in a manner that depended on the neddylated RING sub-complex of the SCF. Taken together, our results identified a new ubiquitin binding activity within the human Cdc34 C terminus that contributes to SCF-dependent ubiquitination.
Figures
References
-
- Hershko A., Ciechanover A. (1998) Annu. Rev. Biochem. 67, 425–479 - PubMed
-
- Pickart C. M. (2004) Cell 116, 181–190 - PubMed
-
- Feldman R. M., Correll C. C., Kaplan K. B., Deshaies R. J. (1997) Cell 91, 221–230 - PubMed
-
- Skowyra D., Craig K. L., Tyers M., Elledge S. J., Harper J. W. (1997) Cell 91, 209–219 - PubMed
-
- Tan P., Fuchs S. Y., Chen A., Wu K., Gomez C., Ronai Z., Pan Z. Q. (1999) Mol. Cell 3, 527–533 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
