

T cells can mediate viral clearance from ependyma but not from brain parenchyma in a major histocompatibility class I- and perforin-independent manner
- PMID: 20354003
- PMCID: PMC7110187
- DOI: 10.1093/brain/awq028
T cells can mediate viral clearance from ependyma but not from brain parenchyma in a major histocompatibility class I- and perforin-independent manner
Abstract
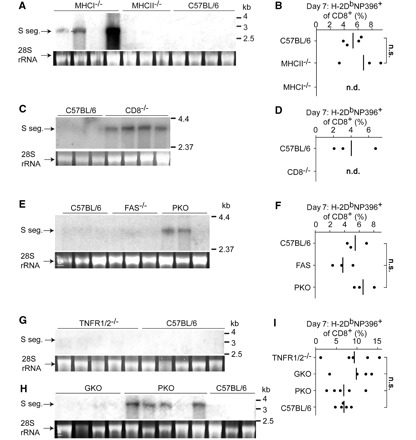
Viral infection of the central nervous system can lead to disability and death. Yet the majority of viral infections with central nervous system involvement resolve with only mild clinical manifestations, if any. This is generally attributed to efficient elimination of the infection from the brain coverings, i.e. the meninges, ependyma and chorioplexus, which are the primary targets of haematogeneous viral spread. How the immune system is able to purge these structures from viral infection with only minimal detrimental effects is still poorly understood. In the present work we studied how an attenuated lymphocytic choriomeningitis virus can be cleared from the central nervous system in the absence of overt disease. We show that elimination of the virus from brain ependyma, but not from brain parenchyma, could be achieved by a T cell-dependent mechanism operating independently of major histocompatibility class I antigens and perforin. Considering that cytotoxic T lymphocyte-mediated cytotoxicity is a leading cause of viral immunopathology and tissue damage, our findings may explain why the most common viral intruders of the central nervous system rarely represent a serious threat to our health.
Figures




Similar articles
-
Protection of CD3 delta knockout mice from lymphocytic choriomeningitis virus-induced immunopathology: implications for viral neuroinvasion.Virology. 2000 Apr 10;269(2):248-56. doi: 10.1006/viro.2000.0224. Virology. 2000. PMID: 10753703
-
The expression of major histocompatibility complex (MHC) class I antigens in the brain differs markedly in acute and persistent infections with lymphocytic choriomeningitis virus (LCMV).J Neuroimmunol. 1992 Feb;36(2-3):193-8. doi: 10.1016/0165-5728(92)90050-u. J Neuroimmunol. 1992. PMID: 1732281 Free PMC article.
-
Thymic tolerance to only one viral protein reduces lymphocytic choriomeningitis virus-induced immunopathology and increases survival in perforin-deficient mice.J Virol. 1999 Jul;73(7):5918-25. doi: 10.1128/JVI.73.7.5918-5925.1999. J Virol. 1999. PMID: 10364344 Free PMC article.
-
Viral escape from the neutralizing antibody response: the lymphocytic choriomeningitis virus model.Immunogenetics. 2001 Apr;53(3):185-9. doi: 10.1007/s002510100314. Immunogenetics. 2001. PMID: 11398962 Review.
-
A suspenseful game of 'hide and seek' between virus and host.Nat Immunol. 2007 Apr;8(4):325-7. doi: 10.1038/ni0407-325. Nat Immunol. 2007. PMID: 17375090 Review.
Cited by
-
Brain-resident memory T cells represent an autonomous cytotoxic barrier to viral infection.J Exp Med. 2016 Jul 25;213(8):1571-87. doi: 10.1084/jem.20151916. Epub 2016 Jul 4. J Exp Med. 2016. PMID: 27377586 Free PMC article.
-
Neuroprotective intervention by interferon-γ blockade prevents CD8+ T cell-mediated dendrite and synapse loss.J Exp Med. 2013 Sep 23;210(10):2087-103. doi: 10.1084/jem.20122143. Epub 2013 Sep 2. J Exp Med. 2013. PMID: 23999498 Free PMC article.
-
Multiciliated ependymal cells: an update on biology and pathology in the adult brain.Acta Neuropathol. 2024 Sep 10;148(1):39. doi: 10.1007/s00401-024-02784-0. Acta Neuropathol. 2024. PMID: 39254862 Review.
-
Antigen recognition detains CD8+ T cells at the blood-brain barrier and contributes to its breakdown.Nat Commun. 2023 May 30;14(1):3106. doi: 10.1038/s41467-023-38703-2. Nat Commun. 2023. PMID: 37253744 Free PMC article.
-
CXCR4 and CXCR6 dually limit T cell entry into the polyomavirus-infected brain.J Neuroinflammation. 2025 Jun 28;22(1):169. doi: 10.1186/s12974-025-03496-2. J Neuroinflammation. 2025. PMID: 40581668 Free PMC article.
References
-
- Adachi M, Suematsu S, Kondo T, Ogasawara J, Tanaka T, Yoshida N, et al. Targeted mutation in the Fas gene causes hyperplasia in peripheral lymphoid organs and liver. Nat Genet. 1995;11:294–300. - PubMed
-
- Adair CV, Gauld RL, Smadel JE. Aseptic meningitis, a disease of diverse etiology: clinical and etiologic studies on 854 cases. Ann Intern Med. 1953;39:675–704. - PubMed
-
- Ahmed R, Gray D. Immunological memory and protective immunity: understanding their relation. Science. 1996;272:54–60. - PubMed
-
- Andersen IH, Marker O, Thomsen AR. Breakdown of blood-brain barrier function in the murine lymphocytic choriomeningitis virus infection mediated by virus-specific CD8+ T cells. J Neuroimmunol. 1991;31:155–63. - PubMed
-
- Badovinac VP, Porter BB, Harty JT. Programmed contraction of CD8(+) T cells after infection. Nat Immunol. 2002;3:619–26. - PubMed
