

Identifying physiological origins of baroreflex dysfunction in salt-sensitive hypertension in the Dahl SS rat
- PMID: 20354102
- PMCID: PMC2888563
- DOI: 10.1152/physiolgenomics.00027.2010
Identifying physiological origins of baroreflex dysfunction in salt-sensitive hypertension in the Dahl SS rat
Abstract
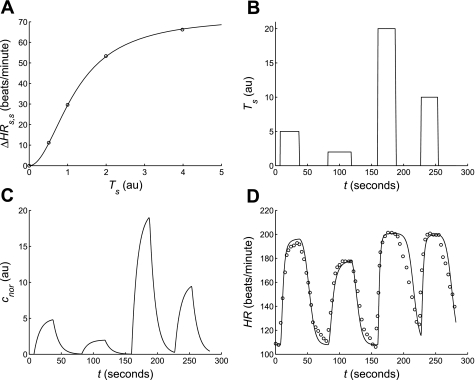
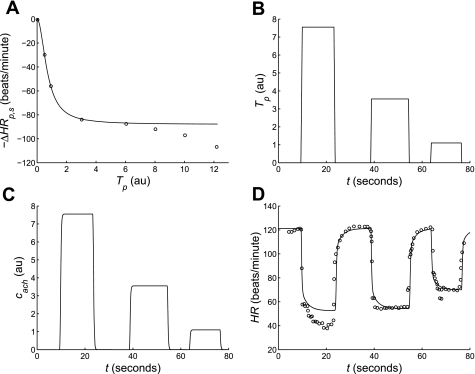
Salt-sensitive hypertension is known to be associated with dysfunction of the baroreflex control system in the Dahl salt-sensitive (SS) rat. However, neither the physiological mechanisms nor the genomic regions underlying the baroreflex dysfunction seen in this rat model are definitively known. Here, we have adopted a mathematical modeling approach to investigate the physiological and genetic origins of baroreflex dysfunction in the Dahl SS rat. We have developed a computational model of the overall baroreflex heart rate control system based on known physiological mechanisms to analyze telemetry-based blood pressure and heart rate data from two genetic strains of rat, the SS and consomic SS.13(BN), on low- and high-salt diets. With this approach, physiological parameters are estimated, unmeasured physiological variables related to the baroreflex control system are predicted, and differences in these quantities between the two strains of rat on low- and high-salt diets are detected. Specific findings include: a significant selective impairment in sympathetic gain with high-salt diet in SS rats and a protection from this impairment in SS.13(BN) rats, elevated sympathetic and parasympathetic offsets with high-salt diet in both strains, and an elevated sympathetic tone with high-salt diet in SS but not SS.13(BN) rats. In conclusion, we have associated several important physiological parameters of the baroreflex control system with chromosome 13 and have begun to identify possible physiological mechanisms underlying baroreflex impairment and hypertension in the Dahl SS rat that may be further explored in future experimental and modeling-based investigation.
Figures










Similar articles
-
Baroreflex control of heart rate in young and adult salt hypertensive inbred Dahl rats.Physiol Res. 2000;49(3):323-30. Physiol Res. 2000. PMID: 11043919
-
Central and baroreflex control of heart period during the wake-sleep cycle in consomic rats with different genetic susceptibility to hypertension.Clin Exp Pharmacol Physiol. 2010 Mar;37(3):322-7. doi: 10.1111/j.1440-1681.2009.05293.x. Epub 2009 Sep 21. Clin Exp Pharmacol Physiol. 2010. PMID: 19769608
-
Molecular networks in Dahl salt-sensitive hypertension based on transcriptome analysis of a panel of consomic rats.Physiol Genomics. 2008 Jun 12;34(1):54-64. doi: 10.1152/physiolgenomics.00031.2008. Epub 2008 Apr 22. Physiol Genomics. 2008. PMID: 18430809
-
Role of the CYP4A/20-HETE pathway in vascular dysfunction of the Dahl salt-sensitive rat.Clin Sci (Lond). 2013 Jun;124(12):695-700. doi: 10.1042/CS20120483. Clin Sci (Lond). 2013. PMID: 23438293 Free PMC article. Review.
-
Sympathetic neural contribution to salt-induced hypertension in Dahl rats.Hypertension. 1991 Jan;17(1 Suppl):I86-90. doi: 10.1161/01.hyp.17.1_suppl.i86. Hypertension. 1991. PMID: 1987017 Review.
Cited by
-
Reversal of genetic salt-sensitive hypertension by targeted sympathetic ablation.Hypertension. 2013 Apr;61(4):806-11. doi: 10.1161/HYPERTENSIONAHA.111.00474. Epub 2013 Feb 4. Hypertension. 2013. PMID: 23381790 Free PMC article.
-
Multiscale modeling and data integration in the virtual physiological rat project.Ann Biomed Eng. 2012 Nov;40(11):2365-78. doi: 10.1007/s10439-012-0611-7. Epub 2012 Jul 18. Ann Biomed Eng. 2012. PMID: 22805979 Free PMC article.
-
Expression of Proinflammatory Cytokines Is Upregulated in the Hypothalamic Paraventricular Nucleus of Dahl Salt-Sensitive Hypertensive Rats.Front Physiol. 2018 Feb 22;9:104. doi: 10.3389/fphys.2018.00104. eCollection 2018. Front Physiol. 2018. PMID: 29520237 Free PMC article.
-
Acute and long-term effects of cannabinoids on hypertension and kidney injury.Sci Rep. 2022 Apr 12;12(1):6080. doi: 10.1038/s41598-022-09902-6. Sci Rep. 2022. PMID: 35413977 Free PMC article.
-
Parameter subset selection techniques for problems in mathematical biology.Biol Cybern. 2019 Apr;113(1-2):121-138. doi: 10.1007/s00422-018-0784-8. Epub 2018 Oct 30. Biol Cybern. 2019. PMID: 30377765 Free PMC article.
References
-
- Alfrey KD. A Model of the Aortic Baroreceptor in Rat (MS thesis) Houston, TX: Rice University, 1997
-
- Andresen MC. High-salt diet elevates baroreceptor pressure thresholds in normal and Dahl rats. Circ Res 64: 695–702, 1989 - PubMed
-
- Andresen MC. Short- and long-term determinants of baroreceptor function in aged normotensive and spontaneously hypertensive rats. Circ Res 54: 750–759, 1984 - PubMed
-
- Andresen MC, Rudis SK, Bee DE. Aberrant baroreceptor mechanotransduction in adult Dahl rats on low-salt diet. Am J Physiol Heart Circ Physiol 256: H446–H454, 1989 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical