

Comparative analysis of right element mutant lox sites on recombination efficiency in embryonic stem cells
- PMID: 20356367
- PMCID: PMC2865440
- DOI: 10.1186/1472-6750-10-29
Comparative analysis of right element mutant lox sites on recombination efficiency in embryonic stem cells
Abstract
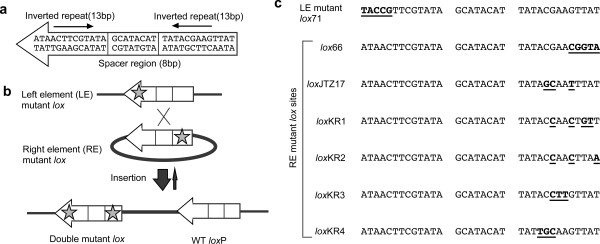
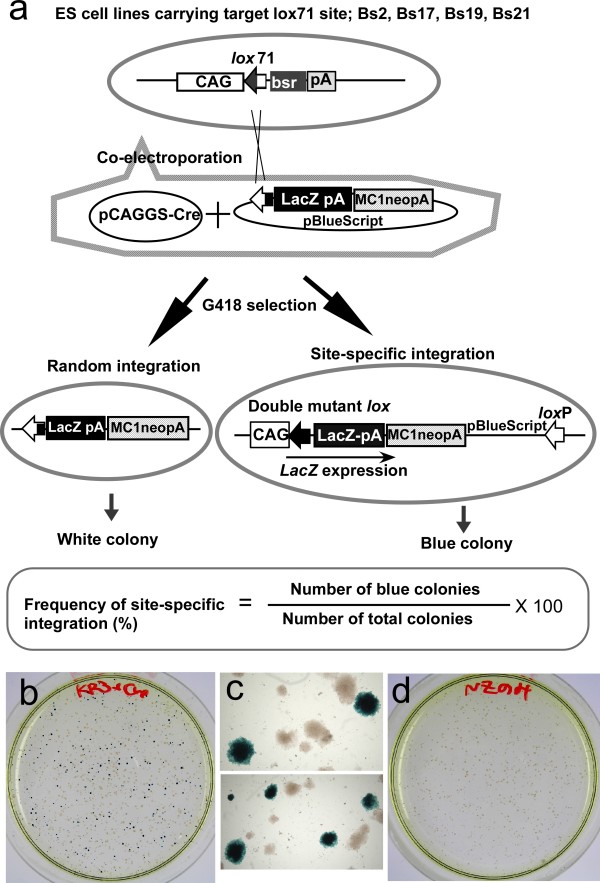
Background: Cre-mediated site-specific integrative recombination in mouse embryonic stem (ES) cells is a useful tool for genome engineering, allowing precise and repeated site-specific integration. To promote the integrative reaction, a left element/right element (LE/RE) mutant strategy using a pair of lox sites with mutations in the LE or RE of the lox sequence has previously been developed. Recombination between LE and RE mutant lox produces a wild-type loxP site as well as an LE+RE double mutant lox site, which has mutations in both sides and less affinity to Cre, resulting in stable integration. We previously demonstrated successful integrative recombination using lox71 (an LE mutant) and lox66 (an RE mutant) in ES cells. Recently, other LE/RE mutant lox sites showing higher recombination efficiency in Escherichia coli have been reported. However, their recombination efficiency in mammalian cells remains to be analyzed.
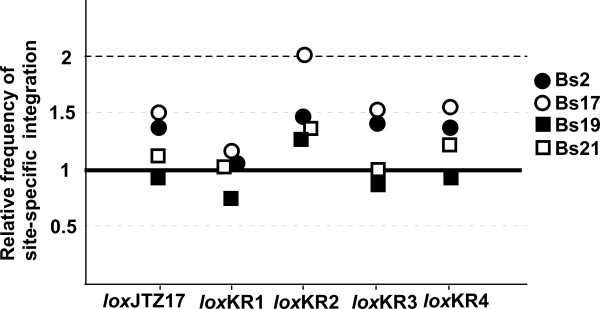
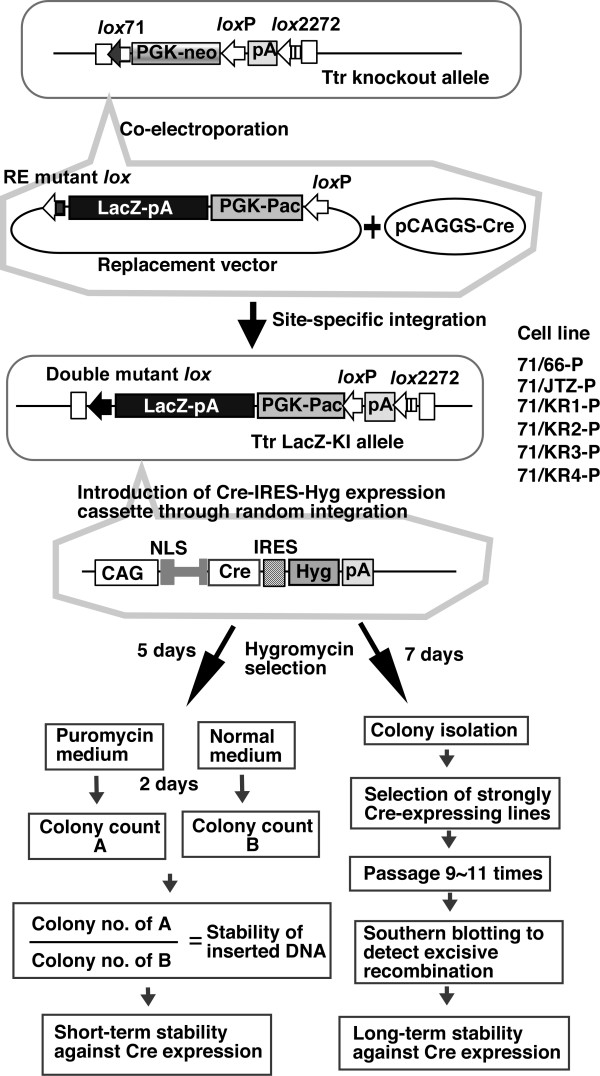
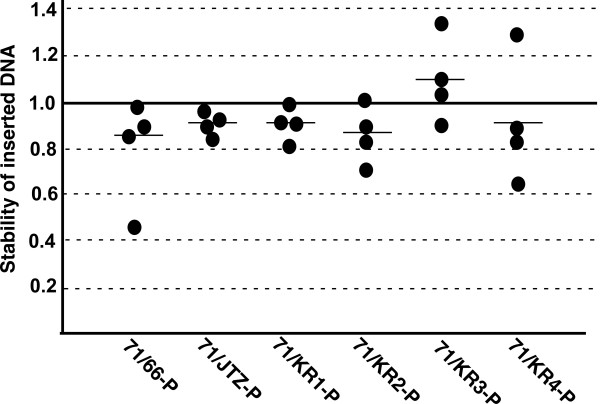
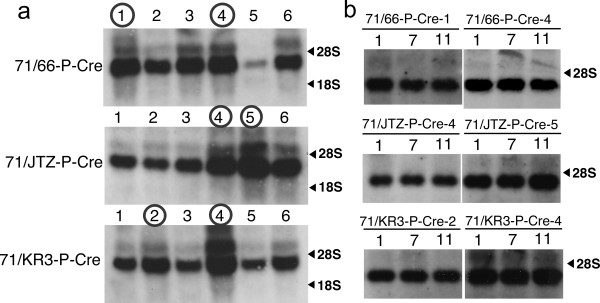
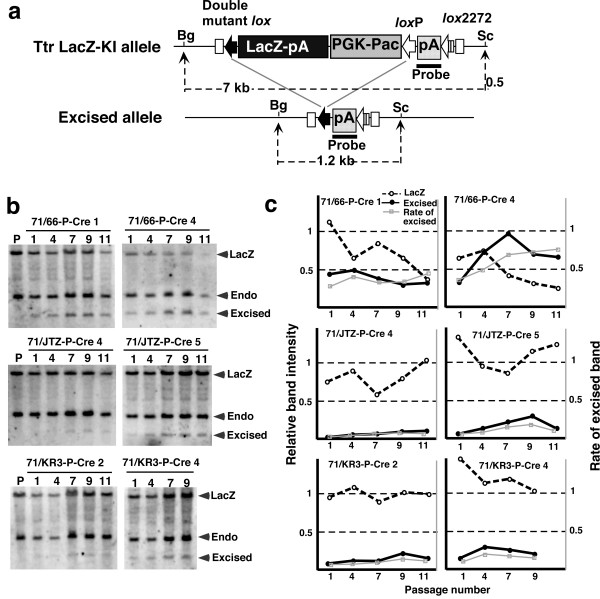
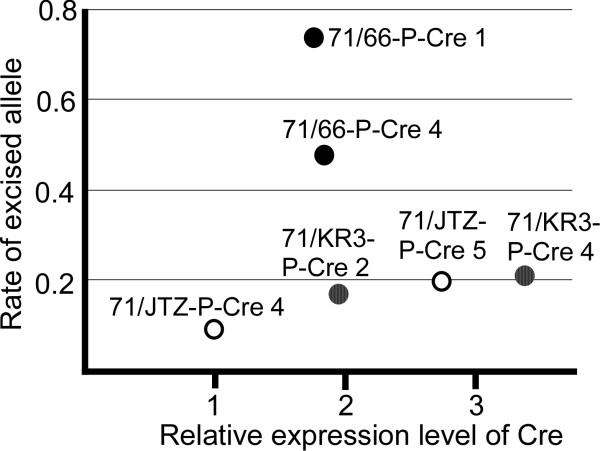
Results: Using ES cells, we compared six RE mutant lox sites, focusing on their recombination efficiency with lox71. All of the RE mutant lox sites showed similar recombination efficiency. We then analyzed the stability of the recombined product, i.e., the LE+RE double mutant lox site, under continuous and strong Cre activity in ES cells. Two RE mutants, loxJTZ17 and loxKR3, produced more stable LE+RE double mutant lox than did the lox66/71 double mutant.
Conclusion: The two mutant RE lox sites, loxJTZ17 and loxKR3, are more suitable than lox66 for Cre-mediated integration or inversion in ES cells.
Figures
Similar articles
-
Site-directed integration of the cre gene mediated by Cre recombinase using a combination of mutant lox sites.Nucleic Acids Res. 2002 Oct 1;30(19):e103. doi: 10.1093/nar/gnf102. Nucleic Acids Res. 2002. PMID: 12364620 Free PMC article.
-
Targeted integration of DNA using mutant lox sites in embryonic stem cells.Nucleic Acids Res. 1997 Feb 15;25(4):868-72. doi: 10.1093/nar/25.4.868. Nucleic Acids Res. 1997. PMID: 9016639 Free PMC article.
-
Site-directed integration system using a combination of mutant lox sites for Corynebacterium glutamicum.Appl Microbiol Biotechnol. 2007 Dec;77(4):871-8. doi: 10.1007/s00253-007-1215-2. Epub 2007 Oct 16. Appl Microbiol Biotechnol. 2007. PMID: 17938910
-
[Progress in Cre/lox site-specific recombination system in higher eukaryotes].Yi Chuan. 2012 Feb;34(2):177-89. doi: 10.3724/sp.j.1005.2012.00177. Yi Chuan. 2012. PMID: 22382059 Review. Chinese.
-
[Commonly used cre transgenic mice and their applications in hematopoietic system].Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2014 Oct;22(5):1442-7. doi: 10.7534/j.issn.1009-2137.2014.05.049. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2014. PMID: 25338604 Review. Chinese.
Cited by
-
Improved tools for the Brainbow toolbox.Nat Methods. 2013 May 5;10(6):540-7. doi: 10.1038/nmeth.2450. Epub 2013 May 5. Nat Methods. 2013. PMID: 23817127 Free PMC article.
-
Testing the Cre-mediated genetic switch for the generation of conditional knock-in mice.PLoS One. 2019 Mar 13;14(3):e0213660. doi: 10.1371/journal.pone.0213660. eCollection 2019. PLoS One. 2019. PMID: 30865697 Free PMC article.
-
A photoactivatable Cre-loxP recombination system for optogenetic genome engineering.Nat Chem Biol. 2016 Dec;12(12):1059-1064. doi: 10.1038/nchembio.2205. Epub 2016 Oct 10. Nat Chem Biol. 2016. PMID: 27723747
-
Gene stacking by recombinases.Plant Biotechnol J. 2016 Feb;14(2):471-82. doi: 10.1111/pbi.12459. Epub 2015 Aug 30. Plant Biotechnol J. 2016. PMID: 26332944 Free PMC article. Review.
-
The kick-in system: a novel rapid knock-in strategy.PLoS One. 2014 Feb 19;9(2):e88549. doi: 10.1371/journal.pone.0088549. eCollection 2014. PLoS One. 2014. PMID: 24586341 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
