

Mechanosensitivity of ion channels based on protein-lipid interactions
- PMID: 20356872
- PMCID: PMC2943882
- DOI: 10.1098/rsif.2010.0095.focus
Mechanosensitivity of ion channels based on protein-lipid interactions
Abstract
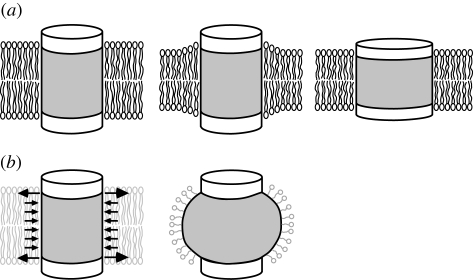
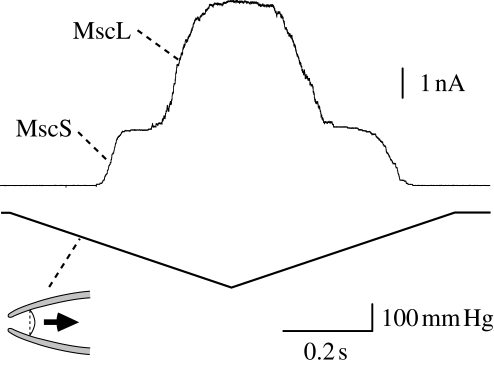
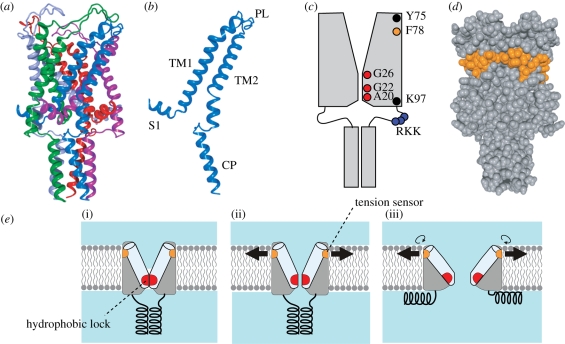
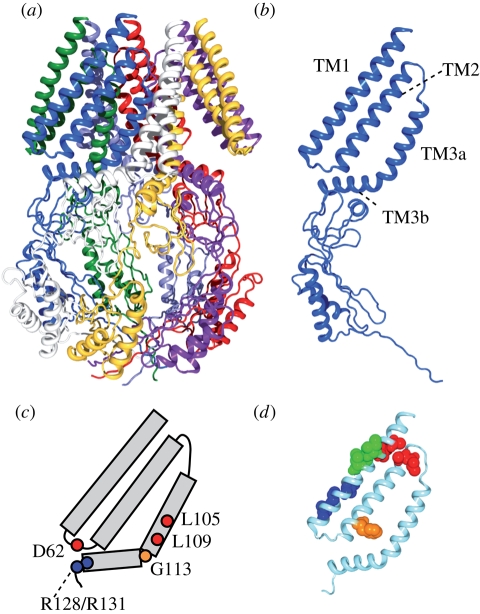
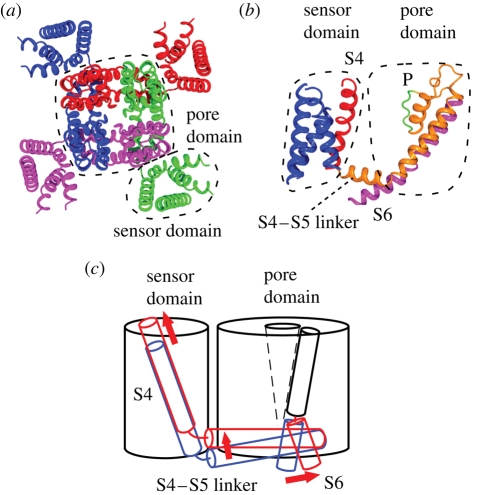
Ion channels form a group of membrane proteins that pass ions through a pore beyond the energy barrier of the lipid bilayer. The structure of the transmembrane segment of membrane proteins is influenced by the charges and the hydrophobicity of the surrounding lipids and the pressure on its surface. A mechanosensitive channel is specifically designed to change its conformation in response to changes in the membrane pressure (tension). However, mechanosensitive channels are not the only group that is sensitive to the physical environment of the membrane: voltage-gated channels are also amenable to the lipid environment. In this article, we review the structure and gating mechanisms of the mechanosensitive channels and voltage-gated channels and discuss how their functions are affected by the physical properties of the lipid bilayer.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources